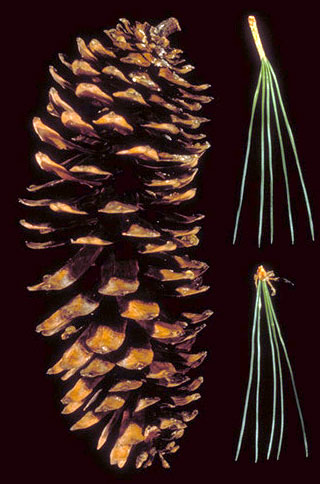
Pinus (Pine)
Linnaeus 1753, p. 1000
Common names
Pine, pinyon [English], Бор [Bulgarian], fyr [Danish], pijn, den [Dutch], mänty [Finnish], pin or pignon [French], Kiefer [German], Πεύκος [Greek], pino [Italian], furu [Norwegian], pinheiro [Portugese], кедр or cосна́ [Russian], bora or molike [Serbo-croat], piñon or pino [Spanish], tall [Swedish], çam [Turkish], chir or kail [Hindi], thong [Vietnamese], 松属 matsu [Japanese], 松属 song shu [Chinese]. Besides these, there are numerous regional names and names in indigenous languages. No language appears to differentiate the subgenera of Pinus, but several languages, such as Russian and Spanish, apply different names to nut pines than to winged-seed pines. The genus name Pinus was the Roman name for pine, and in turn is thought to be derived from an Indo-European root pīt, meaning "resin".
Taxonomic notes
Syn: Apinus Necker; Strobus Opiz; Caryopitys Small; Ducampopinus A. Cheval. (Farjon 1998).
The following table lists all subgenera, sections, subsections, and species of pines.
| Subgenus |
Section |
Subsection |
Link to Species; Notes |
| Pinus |
Pinus |
Pinus |
The classic Old World, 2-needle hard pines:
P. densata,
P. densiflora,
P. henryi,
P. hwangshanensis,
P. kesiya,
P. latteri,
P. luchuensis,
P. massoniana,
P. merkusii,
P. mugo,
P. nigra,
P. resinosa,
P. sylvestris,
P. tabuliformis,
P. taiwanensis,
P. thunbergii,
P. tropicalis (of the Caribbean),
P. uncinata, and
P. yunnanensis.
|
| Pinaster Mayr ex Koehne |
A mostly Mediterranean group: P. brutia,
P. canariensis,
P. halepensis,
P. heldreichii,
P. pinaster,
P. pinea, and
P. roxburghii (of the Himalaya).
|
| Trifoliae Duhamel |
Attenuatae Van Der Burgh |
The fire-adapted, closed-cone pines of California and neighboring areas: P. attenuata,
P. muricata, and
P. radiata.
|
| Australes Loudon |
Based mainly on the analysis of Gernandt et al. (2018) and Cruz-Nicolás et al. (2024), the group includes 5 well-supported clades:
(1) Taeda in the SE US includes
P. echinata,
P. glabra,
P. pungens,
P. rigida,
P. serotina, and
P. taeda; relations within this clade remain unclear.
(2) Elliottii in the SE US and Caribbean includes
the Caribbean species (P. caribaea,
P. cubensis, and
P. occidentalis), sister to
(P. elliottii plus
P. palustris).
(3) Patula in the Sierra Madre Oriental and S to Nicaragua includes
P. greggii,
P. patula, and
P. tecunumanii.
(4) Chihuahuana, mostly in the Sierra Madre Occidental, includes
P. leiophylla sister to the group
P. herrerae,
P. lumholtzii, and
P. teocote.
(5) Oocarpa, mostly in the Sierra Madre Occidental and S to Honduras, includes
P. jaliscana sister to
P. vallartensis; the remaining species include
P. georginae,
P. lawsonii,
P. luzmariae,
P. oocarpa,
P. praetermissa, and
P. pringlei, but relationships between these remain unclear.
|
| Ponderosae Louden |
These pines of the W U.S. and Mexico are sister to subsection Sabinianae, and include
P. arizonica,
P. brachyptera,
P. cooperi,
P. devoniana,
P. durangensis,
P. engelmannii,
P. gordoniana,
P. hartwegii,
P. maximinoi,
P. montezumae,
P. ponderosa,
P. pseudostrobus,
P. scopulorum,
P. stormiae, and
P. yecorensis. See P. ponderosa for an introduction to this subsection.
|
| Sabinianae Louden |
Pines of the W U.S. and Mexico, with
P. jeffreyi sister to a clade comprised of
P. coulteri,
P. sabiniana, and
P. torreyana.
|
| Contortae Little et Critchfield |
North America, including
P. clausa,
P. virginiana, and the
P. banksiana-P. contorta complex.
|
| Strobus Lemmon |
Quinquefoliae Duhamel |
Gerardianae Loudon |
A group of unusual E Asian pines: P. bungeana,
P. gerardiana, and
P. squamata.
|
| Krempfianae Little et Critchfield |
A unique Vietnamese pine, P. krempfii.
|
| Strobus Loudon |
The holarctic nut pines:
P. albicaulis,
P. cembra,
P. koraiensis,
P. pumila, and
P. sibirica.
The white pines of East Asia:
P. amamiana,
P. armandii,
P. bhutanica,
P. cernua,
P. dalatensis,
P. fenzeliana,
P. kwangtungensis,
P. morrisonicola,
P. parviflora,
P. ravii,
P. wallichiana, and
P. wangii. (See also Phylogeny of East Asian white pines).
The white pines of North America:
P. ayacahuite,
P. chiapensis,
P. flexilis,
P. lambertiana,
P. monticola,
P. strobiformis,
P. strobus,
P. stylesii, and
P. veitchii.
The unique European pine P. peuce.
|
| Parrya Mayr |
Nelsoniae Van Der Burgh |
The unique Mexican pine P. nelsonii.
|
| Balfourianae Engelmann |
The ancient "foxtail" pines of the SW U.S.: P. aristata,
P. balfouriana, and
P. longaeva.
|
| Rzedowskiae Carvajal |
The "big cone" clade of piñon pines:
P. maximartinezii,
P. pinceana, and
P. rzedowskii.
|
| Cembroides Engelmann |
The "small cone" clade of piñon pines:
the "cembroides" group P. cembroides,
P. culminicola,
P. discolor,
P. johannis,
P. lagunae,
and
P. remota; and
the "monophylla" group P. californiarum,
P. edulis,
P. monophylla, and
P. quadrifolia.
|
The pine genus is the largest in the Pinophyta, with 126 species treated here, plus quite a few subspecies, varieties, and named hybrids. Pinus is one of the oldest extant conifer genera. The oldest pine known is Pinus mundayi, a 2-needle pine described from fossil charcoal of the lower Cretaceous (ca. 133–140 Ma) Chaswood Formation of Nova Scotia, Canada (Falcon-Lang et al. 2016); the charcoal occurrence of this fossil is interesting, as it suggests that this extremely fire-adapted genus may have had an association with fire since the beginning. A comparable ancient fossil pine is Pinus yorkshirensis, documented from a fossil cone preserved in the Wealden Formation in Yorkshire, UK; these sediments are about 130 million years old (Ryberg et al. 2012).
Pines include two subgenera, the hard pines in subgenus Pinus and the white pines in subgenus Strobus. Some authorities also designate a third subgenus Ducampopinus with species that in the traditional classification are assigned to Strobus section Parrya. Each of the subgenera has been treated as a genus in its own right, and other subgenera have been proposed, but the great majority of morphological evidence, terpene data, and, more recently, molecular phylogenetic data have all firmly established the species composition and monophyletic origin of these two subgenera and the major sections within them. The genetic distance between the subgenera may be as large as or larger than that between e.g. Keteleeria Carrière and Abies Miller (Price et al. 1987), and if strict genetic criteria were used they should perhaps be treated at generic rank. However, the pines as a whole form a clear monophyletic unit and retention as a single genus remains an appropriate treatment. The nomenclatural upheaval of splitting the genus would also be a serious problem.
The subgenera are chiefly distinguished by the following criteria (Richardson 1998):
Subgenus Pinus L., also called the hard pines. About 74 species. Cone scales have a dorsal, mucronate umbo; scales have a sealing band adjacent to the apophysis where the scales meet on the closed cone. There are two leaf vascular bundles per needle. Fascicles have 2-6 needles, stomata are more or less equally distributed on all surfaces, resin ducts are variable; sheath is persistent except in P. leiophylla and P. lumholtzii; the fascicle bases have decurrent pulvini.
Subgenus Strobus Lemmon (syn. genera Apinus Necker; Caryopitys Small; Ducampopinus A. Cheval.; Strobus Opiz), also called white or soft pines. About 45 species. Cone scales lack a sealing band; seed wing is articulate to strongly adnate. There is one leaf vascular bundle per needle. Fascicles have 1-5 needles, stomata are all or mostly on inner faces, resin ducts are medial or external; the fascicle sheath is deciduous except in P. nelsonii, and the fascicle bases have non-decurrent pulvini.
Each of the subgenera can be further subdivided into a number of sections and subsections (listed in the above table). There is a long history of disagreement on the species composition of most of the sections and subsections, although of course some closely related species or groups of species have been long and widely recognized (e.g. the closed-cone pines), and molecular analyses performed mostly in the 21st century have largely ended the disagreements, although there remain some significant problems with molecular analyses of Pinus (Syring et al. 2007, Willyard et al. 2009) which continue to confound understanding of large subsections such as Ponderosae. The sections and subsections presented here are mainly those of Gernandt et al. (2005) (amended to include subsections Attenuatae and Sabinianae) and are based on their cladistic analysis using both character state and molecular phylogenetic data, as well as on molecular phylogenetic studies published by Liston et al. (1999), Wang et al. (1999), Geada López et al. (2002), Syring et al. (2005, 2007), Hernández-León et al. (2013), Jin et al. (2021), Willyard et al. (2021), and Montes et al. (2022).
Data collected thus far support the idea that the two subgenera are valid, clearly distinct, and are each monophyletic. These data also indicate that subgenus Strobus is probably of earlier derivation, with the species in section Parrya (particularly P. nelsonii) the most primitive in the genus. Each of the sections appears to be monophyletic, with very ancient divisions drawn between the subsections. There are also groups within some the subsections that appear to be monophyletic (as noted in the table above). However, Pinus is one conifer genus that, despite its antiquity, shows ample evidence of ongoing speciation, and this makes it a very taxonomically complex group; see, e.g., Syring et al. (2007) and Willyard et al. (2009, 2021) for discussion of some of the ongoing evolutionary processes producing this complexity.
Description
Trees or shrubs, aromatic, evergreen; crown usually conic when young, often rounded or flat-topped with age. Bark of older stems variously furrowed and plated, plates and/or ridges layered or scaly. Branches usually in pseudowhorls; shoots dimorphic with long shoots and dwarf shoots; dwarf shoots borne in close spirals from axils of scaly bracts and bearing fascicles of leaves (needles). Branchlets stout, ending in a compound bud with many bud scales. Buds ovoid to cylindric, apex pointed (blunt), usually resinous. Leaves dimorphic, spirally arranged; foliage leaves (needles) (1)2-5(6) per fascicle, persisting 2-12 or more years, terete or ± 2-3-angled and rounded on abaxial surface, sessile, sheathed at base by 12-15 overlapping scale leaves, these (at least firmer basal ones) persisting for life of fascicle or shed after first season; resin canals mostly 2 or more (rarely 0-1; max. c. 20). Plants monoecious (rarely semi-dioecious); staminate cones numerous and small, in a dense, spikelike cluster around base of current year's growth, mostly ovoid to cylindric-conic, tan to yellow, red, blue, or lavender. Ovulate cones solitary to few, maturing in 1.5-2(-3) years, shed early or variously persistent, pendent to ± erect, at maturity conic or cylindric, sessile or stalked, shedding seed soon after maturity or variously serotinous (not opening upon maturity but much later, usually in response to fire); scales numerous, persistent, woody or pliable, surface of exposed apical portion of each scale (apophysis) thickened, with umbo (exposed scale surface of young cone) represented by a scar (sometimes apiculate) or extended into a hook, spur, claw, or prickle; bracts included. Two seeds at the base of the cone scale, winged, in some the wing vestigial; cotyledons (3)6-14(24). x=12 (Kral 1993, Little 1980).
Pine anatomy differs from other conifers in several respects, and the resulting anatomical differences are helpful in identification. Here are the major points:
Cones: Cones take two or three years to mature. The time requirement varies between species. Young cones may have a distinctive color, but normally identification requires mature cones. Cones may have relatively stiff, woody scales (the norm in subgenus Pinus) or more flexible scales (subgenus Strobus). The portion of the cone scale that is exposed before the mature cone opens is thickened and is called the umbo; it may be unarmed, or armed with a spine or prickle; and it may be formed into a woody pyramid called an apophysis. The shape and texture of the apophysis varies between species and is an important character for identification. Cones scales may remain closed and sealed by resin long after cone maturity; such cones are adapted to open when the resin is melted by fire and are called serotinous. Normally, cones open at maturity and release seed. The seed may be wind-carried, in which case it is normally small and light with a wing longer than the seed; or it may be dispersed by animals (primarily birds of the family Corvidae, jays and their allies), in which case the seed is normally larger and heavier, and the wing may be reduced to inconspicuity.
Foliage: Pine foliage is of four types: cotyledons, primary leaves, cataphylls and needles. Cotyledons are the first leaves produced when the plant emerges from the seeds. There may be from 4 to 24 of them (24 cotyledons, from Pinus maximartinezii, is the largest number known in any plant). Primary leaves are single, alternate (usually helically arranged), acicular leaves that are usually produced only for the first year of growth but that may be produced for many years in some species (such as Pinus quadrifolia) or may be produced on a mature plant in response to a wound. Cataphylls are alternate (helically arranged) non-chlorophyllous primary leaves produced on shoots; they are typically small, subulate or lanceolate, with erose-hyaline to ciliate margins, leaving a distinctive pattern when they fall off the shoot; they are often a useful character in identification. Needles, of course, are the most common pine leaves. They are borne on dwarf shoots axillary to cataphylls in clusters or fascicles of one to eight needles, initially bound together by a basal sheath that may then fall off or may persist, falling with the needles. The number of leaves per fascicle, the length of needles, the number of sides of the needles (only Pinus monophylla has a round needle), the distribution of stomata (waxy white specks on the leaf surface), and the color and stiffness of the needles can all be useful characters for identification. The interior structure of the leaf also may be important for identification, but this requires a microscope and so leaf anatomical characters are rarely used by field workers.
Bark: Bark characters are usually not too useful for pine identification except after a species has been learned thoroughly in the field. The reason is that many species bear similar bark, and the bark characters change with the age of the tree. However, some species have highly distinctive bark, so it is occasionally an important character in identification. Generally, I provide bark characters for mature trees. Important points to note include the bark color, the size and pattern of fissures in the bark, and whether the bark is scaling or flaking.
See Farjon and Styles (1997) for a much more thorough discussion of pine anatomy.
Volume 1 of the Flora of North America (Kral 1993) offers the following advice to those attempting to identify pine specimens:
- Stated leaf measurements are based on healthy, fully expanded growth, especially that of cone-bearing branches.
- Fascicle-sheath measurements are based on fully developed, unbroken sheaths, not on sheaths as they later break up.
- After pollen is shed, pollen cones may lengthen considerably. Measurements given for pollen cones are those of the cones at the time that pollen is released.
- Colors of seed cones are those of mature, closed or newly opened cones, not of old, open, persistent cones or of weathered serotinous cones. Mature, open cones may be hygroscopic, closing partially or completely when wet.
- Descriptions of apophyses are based on mature, closed or newly opened cones. Unlike characters of umbos of most species, characters of apophyses are much altered as the cone grows.
Distribution and Ecology
Native to all continents and some oceanic islands of the northern hemisphere, chiefly in boreal, temperate, or mountainous tropical regions; reaching its southernmost distribution shortly below the Equator in southeast Asia (Sumatra; P. merkusii). Introduced as ornamental and timber trees in much of the southern hemisphere (Mirov 1967, Kral 1993).
Worldwide distribution of Pinus, generalized to 1x1 degree cells. Red points are native range of taxa; blue points are extralimital occurrences (e.g. agroforestry, horticulture, or naturalized). Sources include data from the BRAHMS database at Conifers of the World, accessed 2021.11.11; and four GBIF Occurrence download on 2021.11.12, DOIs https://doi.org/10.15468/dl.kev9qn, https://doi.org/10.15468/dl.cx89m3, https://doi.org/10.15468/dl.vsd6u3, and https://doi.org/10.15468/dl.ydebe9.
Some of the sections and subsections are geographically quite limited:
Section Quinquefoliae: east Asia, North America and Mexico
Subsection Strobus: east Asia, North America and Mexico
Subsection Krempfianae: Viet Nam
Subsection Gerardianae: China and the Himalaya
Section Parrya: Western United States and Mexico
Subsection Cembroides: Western United States and Mexico
Subsection Balfourianae: Western United States
Subsection Nelsoniae: Mexico
Section Trifoliae: North America, Mexico and the Caribbean
Subsection Attenuatae: Western United States (California) and Mexico (Baja California Norte)
Subsection Australes: United States, Mexico and the Caribbean
Subsection Ponderosae: Western United States and Mexico
Subsection Contortae: North America and Mexico (Baja California Norte)
Section Pinus: Europe, Asia, Mediterreanean Africa, eastern North America and Cuba
Subsection Pinus: Europe, Asia (south to Sumatra), eastern North America and Cuba
Subsection Pinaster: Mediterreanean and western Himalaya
Most pines are fire-adapted, meaning that the recurrence of fire permits pines to maintain a dominant role in forest successions that lead to dominance by non-pines. The precise nature of this fire adaptation varies widely, with some pines tolerant of frequent low-intensity fires and others tending to produce high fuel accumulations that permit stand-destroying fires, after which the pines regenerate quickly. In habitats with infrequent or no fire, pines tend to occur on nutrient-poor sites such as serpentine barrens, lithosols, or bogs. Their low shade tolerance typically precludes growth beneath a closed forest canopy. Many species are very drought tolerant.
In many areas Pinus is a forest dominant, either early successional or longer-lived, persisting in the late successional forest. Certain fire successional species have a "grass stage," i.e., the stem of the young seedling elongates little during the first several years (meanwhile developing a large taproot) and bears many long, curved leaves, the plant then resembling a bunchgrass (Kral 1993). Other species have cones that are long persistent and remain closed, opening only when heated by wildfire (such cones are called serotinous); seeds are released soon after, once the fire is out.
Range maps have been published for the genus, many of its subsections and most of its species by Critchfield and Little (1966).
Remarkable Specimens
Largest and Tallest: For stem volume and diameter, the largest and second-tallest pine species is Pinus lambertiana. The tallest and second-largest pine species is P. ponderosa (subsp. benthamiana), and the third-largest species is P. jeffreyi. All three of these pines reach their greatest size in the mixed conifer forest of the Sierra Nevada mountains of California and the Siskiyou Mountains of SW Oregon. Other species known to attain diameters of greater than 2 meters and proportionate heights include P. bhutanica (2nd largest white pine), P. brutia, P. canariensis (largest pine in the Old World), and P. radiata. Historically, P. lambertiana was tallest and largest by a wide margin, but logging and blister rust have decimated this formerly great tree. Also, historically, P. ponderosa and P. jeffreyi have traded places for second-largest. There is an ancient record of a Pinus nigra 500 cm diameter measured by Alexander the Great, but perhaps that is apocryphal.
Smallest: P. contorta subsp. contorta var. bolanderi grows on extremely nutrient-poor soils and has been known to bear cones when only 20 cm tall; however on good sites it can become a fairly large tree. The smallest maximum size is found in P. culminicola, which is not known to exceed 5 m tall and always grows as a shrub. P. pumila and P. mugo are nearly as small. A wide variety of other species may also be less than a meter tall when found growing at the alpine timberline.
Oldest: Pinus longaeva, of which many individuals over 4,000 years old are known. Due to their occurrence in some very cold and dry environments where disease and stand-destroying disturbance are rare, pines include some extremely long-lived conifers. Ages of over 1000 years have also been encountered in P. albicaulis, P. aristata, P. balfouriana, and P. flexilis, all species native to western North America; and in P. heldreichii of Italy. The last is also the oldest hard pine, and the oldest Old World pine.
Largest natural distribution: P. sylvestris, found from Scotland to Kamchatka, and widely naturalized in the United States and Canada.
Smallest natural distribution: A number of species are severely endangered and have recently experienced large reductions due to human activity, mostly habitat loss. Data on these species is usually old enough that they have likely experienced further declines. Based on available data, the most restricted species is P. squamata, known from a single location in China with 18-20 mature trees. The second-most-restricted species is P. culminicola of Mexico, with a total area of occupancy of something less than 10 km², all on mountaintops where its survival is threatened by climate change and fire.
Highest elevation: P. hartwegii of Mexico is the only species found at elevations above 4000 m, reaching a maximum of 4389 m on the volcano, Nevada de Toluca (photo at right). P. wallichiana and P. contorta subsp. latifolia reach 3900 m, while P. albicaulis, P. culminicola, and P. flexilis all reach 3700 m. The greatest elevation range is found in P. contorta subsp. latifolia, which in some areas occurs less than 100 m above sea level (subsp. contorta occurs at sea level).
Largest seed cones: Pinus coulteri is the most massive, with specimens reported up to 8 pounds (3629 g; Lanner 1999) and 20 inches (51 cm) long (Peattie 1950). The longest cones are in Pinus lambertiana, reaching 58.2 cm (Guinness Book of World Records, accessed 2023.03.13), with slightly larger records in some of the historical sources. The second-heaviest are in Pinus sabiniana. All three are found in California.
Smallest seed cones: Probably P. contorta subsp. contorta var. bolanderi and P. mugo. Almost any species may produce a very small cone that contains no viable seeds, and it really isn't a cone unless it can fulfill its evolutionary purpose of carrying seeds. The smallest cones that contain viable seed are probably found in stressed trees that have small cones at the best of times; P. contorta grows on terrible soils and P. mugo is common at the alpine timberline.
Longest pine needles: P. palustris. The smallest needles, again, are probably found on very small specimens of P. contorta subsp. contorta var. bolanderi.
Most economically important: P. radiata, the most widely planted plantation forestry tree in the world, and as a result, the most widespread pine in the southern hemisphere.
Ethnobotany
Pines are economically important for their timber, pulp, and resin-based products. When the world was tied together by sail, pines often assumed strategic importance as naval stores, thereby influencing patterns of Western colonialism. They were the first timber resource exploited in much of North America. They have long been a principal source of timber for all purposes, including firewood, construction and woodworking. They continue to be a leading genus in agroforestry production, dominating plantations in the U.K. (P. contorta, P. nigra), New Zealand (P. radiata), Brazil (P. elliottii), and other countries.
Stone pines (P. armandii, P. gerardiana, P. pinea, and the species in subsection Cembroides) have an edible seed gathered by indigenous peoples (and sometimes commercially) and often comprising a major seasonal food source. By many accounts, they are also an aphrodisiac (Santesson 2000; see also the Ethnobotany section for P. pinea). Historically, some peoples would eat the inner bark of pines, particularly in times of great hunger.
Historically, many pines were used to produce gum (now called oleoresin), a sticky semi-fluid yellow or brownish resin. "Pine resin flows on the external surface of a tree after a wound is inflicted to form a protective coat that seals the wound to pathogenic microorganisms and prevents loss of sap. To obtain resin commercially, a tapping cut is made in the pine bark and the resin drops are collected into buckets or bags. The principal products of pine resin are rosin and turpentine oil. The most significant hard resin from a commercial point of view is rosin, which is obtained by distillation of pine resin. Rosin is used in paper glue and soap manufacturing, as a constituent of varnishes and paints, and for coating bows of stringed musical instruments. Oil of turpentine is also produced by pine resin distillation and is used for thinning and dissolving paint and varnish, as well as for shoe polish and sealing wax manufacturing. It also has medical properties and can be used as stimulant, antispasmodic, astringent, diuretic and anti-pathogenic. In the past, crude pine resin had been used in sailing vessels as packing material and for waterproofing" (Moussouris and Regato 1999). In 2025, resin-based products from pines are still in worldwide use, with a market size of $6 billion in 2023, growing at 5.5% annually; the growth is largely a by-product of consumer disenchantment with the fossil fuel industry. The primary products are tall oil, rosin, and turpentine. Tall oil is mainly a byproduct of the paper industry and is used in adhesives, inks, and other chemical products. Rosin is mainly produced by tapping trees for their oleoresin, and is used mostly in adhesives, inks, coatings, and rubber. Turpentine, also produced from oleoresin, is used mainly in pine oil disinfectants, though there remains some traditional use as in paints and natural medicine (Straits Research 2024).
Pines are among the most popular of ornamental conifers, particularly in temperate and cold climates, for which many hardy species exist. Although most species see little or no ormamental use, the catalog of those that do is long. This may be because many pines are slow-growing and develop contorted forms that are pleasing to gardeners. Examples of the type include Pinus sylvestris, P. mugo, P. densiflora and P. contorta. Others tend to the opposite extreme, quickly developing into majestic landscape trees. Examples include white pines such as P. strobus and P. armandii, and many pines in subsections Ponderosae (in the new world) and Pinaster (in the old), such as P. ponderosa, P. coulteri, P. montezumae, P. pinaster, P. brutia, and P. canariensis. For most of these popular ornamental species, various cultivars have been developed to emphasize differences in growth form and foliage.
Pines have seen countless uses in research, many tied to the fact that they almost all produce clear annual rings. The science of dendrochronology originated with observations of Pinus ponderosa rings by astronomer Andrew Ellicott Douglass during travels in northern Arizona in 1904. Ever since then, pines have been preferred subjects for dendrochronology. Douglass used tree rings to date the construction of Anasazi ruins in the American Southwest, many workers have used scores of different pines to reconstruct past variations in climate, geochronologists have used pines to determine changes in the rate of atmospheric carbon-14 production over the past 7,000 years, and the rings of pines have been used to address a wide spectrum of other technical problems; for example, the tree-ring pattern in a pine board proved to be a crucial piece of evidence helping to convict the killer of the kidnapped Lindberg baby in 1923 (before O.J. Simpson, perhaps the most celebrated American criminal trial of the 20th century) (Graham 1997).
Observations
Comparative study of members of the genus is most easily accomplished at large arboreta; some are listed at the Links page. You should also try to visit the "hot spots" for the genus; the principal ones are Mexico, California, and the southeast United States.
Remarks
"Strange that so few ever come to the woods to see how the pine lives and grows and spires, lifting its evergreen arms to the light, to see its perfect success; but most are content to behold it in the shape of many broad boards brought to market, and deem that its true success. But the pine is no more lumber than man is, and to be made into boards and houses is no more its true and highest use than the truest use of a man is to be cut down and made into manure. There is a higher law affecting our relation to pines as well as to men. A pine cut down, a dead pine, is no more a pine than a dead human carcass is a man. Can he who has discovered only some of the values of whalebone and whale oil be said to have discovered the true use of the whale? Can he who slays the elephant for his ivory be said to have "seen the elephant" ? These are petty and accidental uses; just as if a stronger race were to kill us in order to make buttons and flageolets of our bones; for everything may serve a lower as well as a higher use. Every creature is better alive than dead, men and moose and pine-trees, and he who understands it aright will rather preserve its life than destroy it" (Thoreau 1858).
Citations
Cruz-Nicolás, Jorge, Juan Pablo Jaramillo-Correa, and David S. Gernandt. 2024. Stochastic processes and changes in evolutionary rate are associated with diversification in a lineage of tropical hard pines (Pinus). Molecular Phylogenetics and Evolution 192:108011, doi.org/10.1016/j.ympev.2024.108011.
Falcon-Lang, Howard J., Viola Mages, and Margaret Collinson. 2016. The oldest Pinus and its preservation by fire. Geology 44(4):303-306.
Gernandt, David S., G. Geada López, S. O. Garcia and Aaron Liston. 2005. Phylogeny and classification of Pinus. Taxon 54(1):29-42.
Gernandt, David S., Xitlali Aguirre Dugua, Alejandra Vázquez-Lobo, Ann Willyard, Alejandra Moreno Letelier, Jorge A. Pérez de la Rosa, Daniel Piñero, and Aaron Liston. 2018. Multi-Locus phylogenetics, lineage sorting, and reticulation in Pinus subsection Australes. American Journal of Botany 105(4):711–25. doi.org/10.1002/ajb2.1052.
Graham, S.A. 1997. Anatomy of the Lindbergh kidnapping. Journal of Forensic Sciences 42(3):368-377.
Hernández-León, Sergio, David S. Gernandt, Jorge A. Pérez de la Rosa, and Lev Jardón-Barbolla. 2013. Phylogenetic relationships and species delimitation in Pinus section Trifoliae inferrred from plastid DNA. PLOS One DOI: 10.1371/journal.pone.0070501.
Jin, Wei-Tao, David S. Gernandt, Christian Wehenkel, Xiao-Mei Xia, Xiao-Xin Wei, and Xiao-Quan Wang. 2021. Phylogenomic and ecological analyses reveal the spatiotemporal evolution of global pines. Proceedings of the National Academy of Sciences 118(20): e2022302118. doi.org/10.1073/pnas.2022302118.
Liston, A., W. A. Robinson, D. Pinero, and E. R. Alvarez-Buylla. 1999. Phylogenetics of Pinus (Pinaceae) based on nuclear ribosomal DNA internal transcribed spacer region sequences. Molecular Phylogenetics and Evolution 11(1):95-109.
Montes, José‐Rubén, Pablo Peláez, Alejandra Moreno‐Letelier, and David S. Gernandt. 2022. Coalescent‐based species delimitation in North American pinyon pines using low‐copy nuclear genes and plastomes. American Journal of Botany 109(5):706–726. doi.org/10.1002/ajb2.1847.
Ryberg, P. E., G. W. Rothwell, R. A. Stockey, J. Hilton, G. Mapes, and J. B. Riding. 2012. Reconsidering relationships among stem and crown group Pinaceae: oldest record of the genus Pinus from the Early Cretaceous of Yorkshire, United Kingdom. International Journal of Plant Sciences 173(8):917-932.
Santesson, Johan. 2000. Pine nuts as aphrodisiacs. http://www.santesson.com/aphrodis/pine.htm, accessed 2002.04.24, now defunct.
Syring, J., K. Farrell, R. Businsky, R. Cronin, and A. Liston. 2007. Widespread genealogical nonmonophyly in species of Pinus subgenus Strobus. Systematic Biology 56(2):163–181
Thoreau, H. D. 1858. Chesuncook. The Atlantic, June.
Straits Research. 2024. Pine Chemicals Market Size, Share & Trends Analysis Report. https://straitsresearch.com/report/pine-chemicals-market, accessed 2025.03.14.
Willyard, Ann, Richard Cronn, and Aaron Liston. 2009. Reticulate evolution and incomplete lineage sorting among the ponderosa pines. Molecular Phylogenetics and Evolution 52:498–511.
Willyard, Ann, David S. Gernandt, Blake Cooper, Connor Douglas, Kristen Finch, Hassan Karemera, Erik Lindberg, Stephen K. Langer, Julia Lefler, Paula Marquardt, Dakota L. Pouncey, and Frank Telewski. 2021. Phylogenomics in the hard pines (Pinus subsection Ponderosae; Pinaceae) confirms paraphyly in Pinus ponderosa, and places Pinus jeffreyi with the California big cone pines. Systematic Botany 46:538–561.
See also
The The Green Pinky provides useful tips on how to plant and grow pines. Their site is a sponsor of the Gymnosperm Database.
Lovett Pinetum, a good source of information on many topics in pine biology.
Axelrod, D. I. 1980. History of the maritime closed-cone pines, Alta and Baja California. Univ. Calif. Publ. Geol. Sci. 120:1-143.
Axelrod, D. I. 1986. Cenozoic history of some western pines. Annals of the Missouri Botanical Garden 73:565-641.
Axelrod, D. I. and J. Cota. 1993. A further contribution to closed-cone pine (Oocarpae) history. American Journal of Botany 80(7):743-751.
Bailey (1970) provides a classic and still very useful treatment of the pines in subsection Balfourianae.
Bailey, D. K. 1987. A study of Pinus subsection Cembroides I: The single-needle Piñons of the Californias and the Great Basin. Notes of the Royal Botanical Gardens Edinburgh 44:275-310.
Bailey, D. K. and F. G. Hawksworth. 1979. Piñons of the Chihuahuan Desert region. Phytologia 44:129-133.
Burns and Honkala (1990) provide detailed descriptions, with a focus on silviculture, for all economically significant pines native to the United States.
Cai, Qing, Daming Zhang, Zhan-Lin Liu and Xiao-Ru Wang. 2006. Chromosomal Localization of 5S and 18S rDNA in Five Species of Subgenus Strobus and their Implications for Genome Evolution of Pinus. Annals of Botany 97(5):715-722.
Critchfield, W. B. 1962. Hybridization of the southern pines in California, pp. 25-27. Proc. Forest Genetics Workshop. Macon, GA.
Critchfield, W. B. 1967. Crossability and relationships of the closed-cone pines. Silvae Genetica 16(3): 89-97.
Critchfield and Little (1969) is still the most encyclopedic source of range maps, though some notable range extensions have since been reported.
Denevan, W. M. 1961. The upland pine forests of Nicaragua. Univ. Cal. Publ. Geol Sci. 12:251-320.
Duffield, J. W. 1952. Relationships and species hybridization in the genus Pinus. Silvae Genetica 1:93-97.
Farjon (1984), or the second edition in 2005, provides a good overview with lots of interesting supplemental information and excellent line drawings.
Farjon, A. 1996. Biodiversity of Pinus (Pinaceae) in Mexico: speciation and paleo-endemism. Botanical Journal of the Linnaean Society 121:365-384.
Farjon and Styles (1997), the most authoritative reference on pines of Mexico and Central America.
Furman, B. J., D. Grattapaglia, W. S. Dvorak, and D. M. O'Malley. 1997. Analysis of genetic relationships of Central American and Mexican pines using RAPD markers that distinguish species. Molecular Ecology 6:321-331.
Gernandt, D. S., S. Magallón, G. Geada López, O. Zerón Flores, A. Willyard, and A. Liston. 2008. Use of simultaneous analyses to guide fossil‐based calibrations of Pinaceae phylogeny. International Journal of Plant Sciences. 169(8):1086–1099.
Hong, Y. P., A. B. Krupkin, and S. H. Strauss. 1993. Chloroplast DNA transgresses species boundaries and evolves at variable rates in the California closed-cone pines (Pinus radiata, P. muricata, and P. attenuata). Genetics 135(4):1187-1196.
Jin, Wei-Tao, David S. Gernandt, Christian Wehenkel, Xiao-Mei Xia, Xiao-Xin Wei, and Xiao-Quan Wang. 2021. Phylogenomic and ecological analyses reveal the spatiotemporal evolution of global pines. Proceedings of the National Academy of Sciences 118(20):e2022302118. doi.org/10.1073/pnas.2022302118.
Krupkin, A. B, A. Liston, and S. H. Strauss. 1996. Phylogenetic analysis of the hard pines (Pinus subgenus Pinus, Pinaceae) from chloroplast DNA restriction site analysis. American Journal of Botany 83(4):489-498.
Liston, A. W., A. Robinson, D. Piñero, and E. R. Alvarez-Buylla. 1999. Phylogenetics of Pinus (Pinaceae) based on nuclear ribosomal DNA internal transcribed spacer region sequences. Molecular Phlyogenetics and Evolution 11(1):95-109.
Little and Critchfield (1969), which established a taxonomic benchmark for the species.
Malusa (1992), a great introduction to the diversity of the piñons.
Millar, C. I. 1993. Impact of the Eocene on the evolution of Pinus L. Annals of the Missouri Botanical Garden 80:471-498.
Millar, C. I., S. H. Strauss, T. H. Conkle, and R. D. Westfall. 1988. Allozyme differentiation and biosystematics of the California closed-cone pines (Pinus subsection Oocarpae). Systematic Botany 13:351-370.
Mirov (1967), for many years the recognized "bible" of pines. Mirov and Hasbrouck (1976), though, offer a more readable introduction to the genus.
Peattie (1950), one of the more 'literate' writers, a man not afraid to confess his love of great trees.
Perry (1991), an invaluable reference for the pines of Mexico and Central America.
Preston, R. J. 1976. North American trees (exclusive of Mexico and tropical United States), ed. 3.
Price, R. A. 1989. The genera of Pinaceae in the southeastern United States. Journal of the Arnold Arboretum 70:247-305.
Price, R. A., A. Liston and S. H. Strauss. 1998. Phylogeny and systematics of Pinus. P.49-68 in Richardson, D.M. (ed.), Ecology and Biogeography of Pinus. Cambridge University Press. ISBN 0-521-55176-5.
Richardson (1998), which provides a wealth of ecological information.
Sargent, C. S. 1922. Manual of the trees of North America (exclusive of Mexico), ed. 2. Boston and New York. [Facsimile edition in 2 vols. 1961, reprinted 1965, New York.]
Shaw (1909, 1914) and Sudworth (1908), both of whom provide excellent drawings, though the taxonomy is inevitably very dated.
Sudworth, G. B. 1917. The pine trees of the Rocky Mountain region. USDA Bulletin 460. Washington, DC.
Syring, J., A. Willyard, R. Cronn, and A. Liston. 2005. Evolutionary relationships among Pinus (Pinaceae) subsections inferred from multiple low-copy nuclear loci. American Journal of Botany 92:2086–2100.
Wu, J. K., V. Krutovskii, and S. H. Strauss. 1999. Nuclear DNA diversity, population differentiation, and phylogenetic relationships in the California closed-cone pines based on RAPD and allozyme markers. Genome 42:893-908.