Conservation Status

Pinus patula
Schiede ex Schlectendahl et Chamisso 1831
Common names
Mexican weeping pine, spreading-leaved pine, patula pine; pino patula, pino chino, pino triste [Spanish] (Wormald 1975).
Taxonomic notes
Two varieties:
1. P. patula Schiede ex Schlechtendal et Chamisso var. patula. Type Mexico, Veracruz: Cruz Blanca, "Inter Cruz blanca & Jalacingo," 1828.11, Schiede and Deppe 1108 (lectotype, HAL). Syn.: Pinus patula Schiede ex Schlechtendal et Chamisso var. macrocarpa M.T. Masters, Gard. Chron., ser. 3, 9: 438, fig. 92. 1891. Type. Gard. Chron., ser. 3, 9: 435, fig. 92. 189 1. (lectotype) (Farjon and Styles 1997).
2. P. patula Schiede ex Schlechtendal et Chamisso var. longipedunculata Loock ex Martínez, Pinos Mexicanos, ed. 2, 333, figs. 276-280, 1948. Type Mexico, Oaxaca: Rancho Benito Juárez (Rancho Tablas), 1947.07, Loock 113a (holotype, PRF-n.v.; isotype, MEXU) (Farjon and Styles 1997).
Farjon and Styles (1997) provide the following notes on the history of the species and its varieties: "Schlechtendal and Chamisso (1831) gave under C.J.W. Schiede & F. Deppe 1108 a concise description of a 3-leaved pine, of which they had seen no cones ('strobilo . . . nobis deficiente') in the material presented to them. ... Gordon (1875) described Pinus patula (var.) macrocarpa based on collections by C.J.W. Schiede with a cone size 'from 6-7 inches long and 2 inches broad', which, if correct, would be quite outside the range of this species as here defined. Masters (1891) formally published this combination and provided an illustration of the cone. No specimen has been traced in the herbaria to match the descriptions and/or the illustration. The illustration in Masters (1891) is therefore designated as the lectotype. ... Martínez (1948) ascribed his new variety P. patula var. longipedunculata to Loock, who coined the name on the label of the type specimen. He described it as different from 'typical’ P. patula in having 2 resin ducts in the leaves instead of 3 and in having pedunculate (5-15 mm) cones versus sessile cones. Perry (1991) allows it occasionally 3 resin ducts."
Farjon and Styles (1997) also report that "Styles (1976) concluded that 'it agrees phenotypically in all botanical characters' with those mentioned by Martinez (1940) under P. oocarpa var. ochoterenae and synonymized both under P. patula. He later expressed a different view (Styles, 1984) but stopped short of formally synonymizing these two taxa under his newly created combination P. patula subsp. tecunumanii. From his determinations of specimens it appears that P. oocarpa var. ochoterenae Martínez was treated as a synonym of P. patula subsp. tecunumanii (see under P. tecunumanii), but that at least more-recently collected material (Higman, Padilla & Styles 40) was again accepted as P. patula (subsp.) longipedunculata. Observation in the field and study of type material by A. Farjon confirm recognition of this taxon as a variety if peduncle length is coupled with persistence and serotiny, which may point to different adaptive traits between the two taxa." Note that Furman et al. (1997) have used RAPD markers to show clear and substantial genetic differences between P. patula and P. tecunumanii, establishing the latter as a good species.
Closely-related species include P. greggii, P. jaliscana, P. oocarpa, P. praetermissa, P. pringlei, and P. tecunumanii, all of which also are native to Mexico.
Description
Trees to 40 m tall and 100 cm dbh, usually with a single, straight, slender trunk; in closed canopy stands, depth of the conical or domed crown depth ca. 33%. Bark on young trees thin, scaly, red-brown, with age becoming thick, dark grey-brown, rough and scaly with large elongated plates and deep longitudinal fissures. First-order branches long, slender, spreading or slightly ascending; higher order branches slender, drooping, the ultimate branches pendent. Shoots rough and scaly when the leaf fascicles have fallen, yellow- to red-brown, foliage shoots with prominent, decurrent pulvini. Cataphylls subulate, recurved at apex, scarious, with erose-ciliate margins, brown, early deciduous. Vegetative buds oblong to cylindrical, the terminal bud 15-20 mm long, the laterals shorter, brown, not resinous; the scales spreading, subulate, ciliate at margins. Fascicle sheaths initially 20-30 mm long with 6-8 imbricate, chartaceous, white-yellow to orange-brown scales, persistent but shortening to 12-15 mm in mature fascicles, slowly weathering to grey-brown. Leaves in fascicles of 3-4(-5), in drooping tufts, typically in two rows on either side of the upturned shoot, persisting 2-3 years, thin, lax, drooping to pendent, (11-)15-25(-30) cm × 0.7-0.9(-1) mm, serrulate at the margins, acute, pale green to dark green. Stomata on all faces of the leaves, in 4-6(-7) lines on the convex abaxial face and 2-3(4) lines on each adaxial face. The leaves are triangular in cross section, the hypodermis usually with a double layer of cells; resin ducts (1-)2-3(4), medial, occasionally 1 internal, stele oval in cross section; outer walls of endodermal cells not thickened; vascular bundles 2, distinct, the xylem strands often connate. Pollen cones crowded near the proximal end of new shoots, spreading, subtended by scarious bracts, ovoid-oblong to cylindrical, 15-20 × 5-6 mm, pink-yellow, turning yellow-brown. Seed cones subterminal or lateral, in whorls of 2 to many, rarely solitary, persistent or deciduous, on short or moderately long (to 20 mm) peduncles. Immature cones ovoid, on short or long recurved peduncles with persistent cataphylls, purple turning light brown, maturing in two seasons. Mature cones narrowly ovoid when closed, usually slightly curved, more or less asymmetrical at base, 5-10(-12) × (3-)4-6.5 cm when open. Seed scales 100-150, serotinous or parting some time after maturing, tenacious or deciduous with the peduncle, (thin) woody, oblong, usually curved when spreading, the proximal scales connate, purple-brown to dark brown, with adaxial light brown marks left by seed wings. Apophysis nearly flat to slightly raised, transversely keeled, more or less gibbous on the proximal scales, more so on one side of the cone, rhombic, upper margin acute or obtuse-rounded, abaxial surface striate or smooth, (lustrous) ochraceous or yellow-brown. Umbo dorsal, flat or raised, often sunken into the apophysis, 3-7 mm wide, grey, with a minute, deciduous prickle. Seeds obliquely ovoid, flattened, 4-6 × 2-4 mm, dark gray. Seed wings articulate, held to the seed by two oblique claws which partly cover the seed on one side, obliquely ovate-oblong, 12-18 × 5-8 mm, light brown with dark stripes (Farjon and Styles 1997). See García Esteban et al. (2004) for a detailed characterization of the wood anatomy.
Pollen dispersal is recorded in February and March (Critchfield, 1966b, cited in Farjon and Styles 1997) (type variety).
Key to the varieties:
Seed cones on (very) short peduncles, seemingly sessile, tenacious, persistent, serotinous, (5-)7-10(-12) cm long, (narrowly) ovoid when closed, broadly ovoid when open. | var. patula |
Seed cones on distinct, curved peduncles, deciduous after a few years, opening readily, 5-8 cm long, narrowly ovoid to ovoid-attenuate when closed, (narrowly) ovoid when open. | var. longepedunculata |
Distribution and Ecology
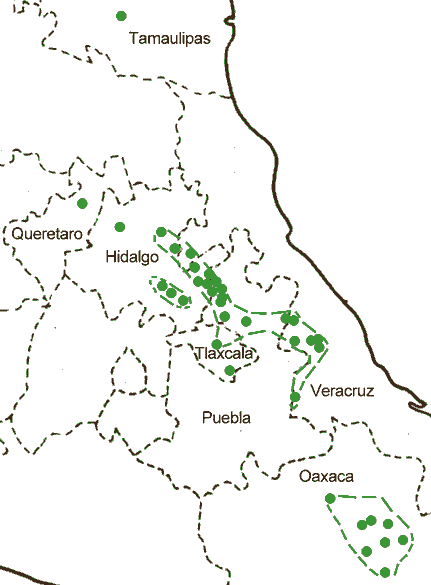
Mexico: Tamaulipas, Querétaro, Hidalgo, México, Distrito Federal, Morelos, Tlaxcala, Puebla, Veracruz, Oaxaca and Chiapas (Farjon and Styles 1997). See also Thompson et al. (1999). This is also a widely-used ornamental and agroforestry species in many moist tropical areas, such as South Africa, hardy to Zone 8 (cold hardiness limit between -12.1°C and -6.7°C) (Bannister and Neuner 2001).
It may occur with a variety of other conifers, particularly
P. devoniana,
P. leiophylla,
P. pseudostrobus,
P. montezumae,
P. lawsonii,
P. ayacahuite,
P. greggii (var. australis),
P. maximinoi,
P. hartwegii, and
P. chiapensis, as well as Taxus globosa,
Podocarpus matudae,
Abies religiosa and
Hesperocyparis lusitanica (Wormald 1975). On drier sites it is commonly replaced by P. teocote, P. montezumae, and P. leiophylla.
The type variety occurs in a few localities in Tamaulipas, in Querétaro, Hidalgo, México, Distrito Federal, Morelos, Tlaxcala, Puebla, Veracruz, Oaxaca, and Chiapas. Its main distribution is on the Sierra Madre Oriental and the high mountains of east-central Mexico, at elevations of (1400-)1500-2800(-3300) m. It is generally restricted to humid, subtropical to warm-temperate sites, with annual precipitation 1000-2200 mm. On the eastern slopes of the Sierra Madre Occidental precipitation is augmented by frequent fog outside the rainy season. Mixed pine-oak forests are most common in the interior, while on the Atlantic slope numerous angiosperms, especially Liquidambar, are (co-)dominant (Farjon and Styles 1997).
Variety longepedunculata is known from two localities in Hidalgo and Veracruz but is more widespread in Oaxaca and central Chiapas; its full distribution is not yet well known. Its ecology and phenology are similar to var. patula, but its ecological amplitude is narrower; it apparently does not occur on the interior mountains of central Mexico, where conditions are drier and seasonal temperature fluctuations more extreme (Farjon and Styles 1997).
Remarkable Specimens
I have no data on exceptional trees in habitat. One specimen near the Redwood Memorial Grove in Rotorua, New Zealand, has been measured at 41.5 m tall (R. Van Pelt email 2009.04.14).
Ethnobotany
This is one of the most important pines for timber in Mexico, as it grows quickly and produces a long, straight, fairly clear bole. It is also widely introduced in other tropical countries in plantation forestry, where in some cases it has become problematic as an invasive species (Farjon and Styles 1997). Dendrochronological research has included work on productivity and wood anatomy done in African plantation forests.
Observations
I have seen notable stands just west of the summit on Mex-175 in Oaxaca (approx. 17.6° N, 96.5° W) and in Querétaro near Pinal del Amoles, at the village of La Cañada (21.117° N, 99.662° W). The species is heavily exploited for timber production at the latter locale. More locations are described by Dvorak et al. (2000) and Perry (1991).
Remarks
The epithet is Latin for "spreading wide" in reference to the leaves, although they are in fact drooping.
This species is one of the principal hosts for the dwarf mistletoes Arceuthobium aureum subsp. petersonii (in Oaxaca and Chiapas), A. globosum subsp. grandicaule (in southern Mexico), A. nigrum (in central and eastern Mexico), and A. vaginatum subsp. vaginatum (widespread in Mexico) (Hawksworth and Wiens 1996).
Citations
Furman, B. J., D. Grattapaglia, W. S. Dvorak and D. M. O’Malley. 1997. Analysis of genetic relationships of Central American and Mexican pines using RAPD markers that distinguish species. Molecular Ecology 6: 321-331. Abstract: Phylogenetic relationships were inferred for six Central American and Mexican pine species by analyzing RAPD marker differences among pooled DNA samples. This population level pooling strategy discounts low-frequency allelic variation within taxa, thus obtaining a ‘cumulative genotype’ to compare among taxa. We used the morphologically based taxonomy of pines as the basis for inference concerning molecular marker divergence. Only RAPD polymorphisms that were repeatable and intensely amplified were used. The resulting data set was found to have strong hierarchical structure. Phylogenetic analyses were carried out using both neighbor-joining and maximum parsimony. The phylogenetic relationships indicated by analysis of the pine RAPD data provide new insights on pine taxonomy. The phylogenetic analyses of the RAPD marker data placed Pinus tecunumanii and P. patula clearly into two different subgroups, strongly supporting the classification of P. tecunumanii as a distinct species, and raise a new set of issues concerning the distinctions between Pinus tecunumanii and P. caribaea. Phylogenetically informative RAPD markers will be useful tools to address ex-situ conservation issues that involve identification and species admixture.
Schlechtendal, D. F. L. von, and A. von Chamisso. 1831. Plantarum mexicanarum a cel. viris Schiede et Deppe collectarum (recensio brevis). Addenda (1). Linnaea 6: 352-384 (p. 354).
Wormald, T.J. 1975. Pinus patula. Tropical Forestry Papers No. 7. Unit of Tropical Silviculture, Department of Forestry, Oxford England.
See also
Dvorak, W.S., J.K. Donahue and J.A. Vásquez. 1995. Early Performance of CAMCORE Introductions of Pinus patula in Brazil, Colombia and South Africa. South African Forestry Journal 174: 23-32. Abstract: Seeds were collected from 13 provenances and 282 mother trees of Pinus patula Schiede & Deppe across a 900 km transect from Tamaulipas to Oaxaca, Mexico by the CAMCORE Cooperative, North Carolina State University, USA. Thirty-three provenance/progeny tests were established in Brazil, Colombia, and South Africa and assessed for height and diameter (DBH) at three years of age. Sources from southern Oaxaca (Cuajimoloyas, Manzanal, and Tlacuache) showed great susceptibility to cold at Jessievale and Commonage, South Africa (26% survival) versus the other P. patula sources (70%) survival). Average height growth at three years of age exceeded 5 m in tests at Los Guadales, Colombia, and Maxwell, South Africa. The best provenance for height growth and volume in both Brazil and South Africa were those from the central part of the P. patula range: Potrero de Monroy (Veracruz), Zacualtipan, (Hidalgo) and Pinal de Amoles (Querétaro). In the Andean mountains of Colombia, the southern Oaxacan sources of P. patula performed as well as the best sources from central Mexico. Some CAMCORE open-pollinated families performed as well as or better in height growth than advanced generation control lots of P. patula, P. taeda or P. elliotti in 29 of 33 tests.
Dvorak, W. S. 1997. The improvement and breeding of Pinus patula, pp. 53-68. In: Proc. 24th South. For. Tree Improv. Conf. Orlando FL.
Dvorak, W.S. 2003. Pinus patula. Species description in the Tropical Tree Seed Manual. Available http://www.rngr.net/Publications/ttsm/Folder.2003-07-11.4726 (accessed 2007.08.31).
Dvorak, W.S., G.R. Hodge, J.E. Kietzka, F. Malan, L.F. Osonio, and T.K. Stanger. 2000. Pinus patula. Pp. 148-173 in Conservation & Testing of Tropical & Subtropical Forest Tree Species by the CAMCORE Cooperative. Raleigh, NC: College of Natural Resources, NCSU.
Elwes and Henry 1906-1913 at the Biodiversity Heritage Library. This series of volumes, privately printed, provides some of the most engaging descriptions of conifers ever published. Although they only treat species cultivated in the U.K. and Ireland, and the taxonomy is a bit dated, still these accounts are thorough, treating such topics as species description, range, varieties, exceptionally old or tall specimens, remarkable trees, and cultivation. Despite being over a century old, they are generally accurate, and are illustrated with some remarkable photographs and lithographs.
The PROTA database account for this species (accessed 2019.02.28). PROTA accounts are focused on commercial forest uses in Africa, and typically include photographs, drawings, names, distribution, and a variety of information relevant to management of the species.