Pinus ponderosa
Douglas ex Lawson 1836
Common names
Ponderosa pine; yellow, western yellow, bull, black Jack, western red, western pitch, big, heavy, Sierra brownbark, or western longleaf pine; pino real, pinabete [Spanish]; pin à bois lourd [French].
Taxonomic notes
After well over a century of taxonomic neglect, during which time almost every cinnamon-barked pine in the western U.S. came to be called "ponderosa pine", the 21st century saw a flurry of increasingly detailed analyses that has now led to the identification of at least five distinct taxa, as well as illuminating relationships within subsection Ponderosae in general. The following discussion therefore addresses all taxa in the subsection, viz.:
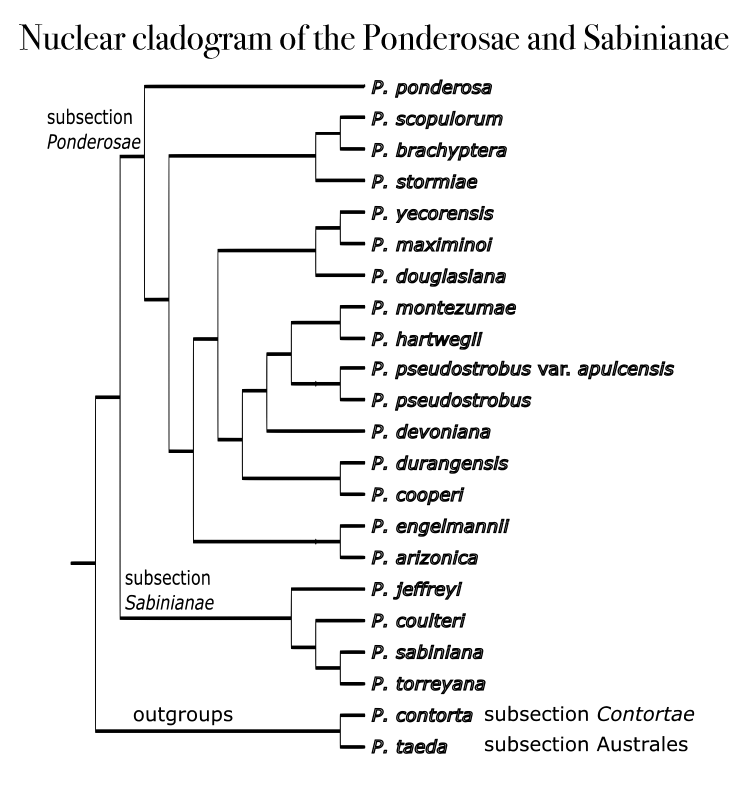
The cladogram at right summarizes current thinking on phylogenetic relationships within subsection Ponderosae (and the closely-related subsection Sabinianae) as inferred on the basis of nuclear genetic data (Willyard et al. 2021b). Other lines of evidence, including plastid and mitochondrial data, generally support this phylogeny (Potter et al. 2013, Willyard et al. 2021a). This cladogram is also generally supported by experimental hybridization results (few of these taxa hybridize in nature). Experiments in cross-pollination among 12 taxa of subsection Ponderosae generally yielded fertile seeds, thus demonstrating their potential to hybridize (Conkle and Critchfield 1988; Critchfield 1966); in addition, P. jeffreyi crosses with P. ponderosa in an uncommon instance of hybridization between two different subsections of Pinus. Many relationships within the cladogram are surprising, though, in the context of traditional morphologically-based interpretations of subsection Ponderosae. Most notably, the Pacific, Washoe, and North Plateau taxa (formerly called "races") of P. ponderosa are polyphyletic with respect to the Rocky Mountain and Southwestern races, now segregated as P. brachyptera and P. scopulorum. Those two species are sister to P. arizonica var. stormiae which has a generally disjunct distribution and has only been described as a variety of either P. arizonica or P. ponderosa, each of which is now seen to be quite a distant relative. It should be described as a species, P. stormiae, but that has not yet occurred. P. douglasiana, P. maximinoi, and P. yecorensis are related, which is morphologically surprising although biogeographically plausible, as they share a primary distribution within the Sierra Madre Occidental. The clade that includes P. cooperi, P. devoniana, P. durangensis, P. hartwegii, P. montezumae, and P. pseudostrobus is plausible, but it is important to note that genetic sampling has been rather light within the distributions of these species and it is entirely possible that their detailed relationships will be understood differently as further information becomes available. It's noteworthy, though, that this analysis has high confidence that P. cooperi, formerly treated as a variety of P. arizonica, is a distinct species. Finally, P. engelmannii and P. arizonica are allied, a somewhat surprising result given their strong morphological differences.
In reviewing the phylogeny of conifers, we don't always appreciate that because they are dominated by relict species, they give an artificially tidy view of evolutionary relationships, so that a cladogram can be seen as a reasonably accurate portrayal of phylogenetic relationships. This is definitely NOT the case in subsection Ponderosae. In this group, review of different molecular markers (e.g., nuclear, chloroplast, and mitochondrial nucleic acids) often indicates that past events have not featured simple dichotomies of evolution. There have been gene transference events, hybridization events, episodes of introgression that lasted for some time and then ended, and times of incomplete lineage sorting (meaning that evolution is happening too quickly for the affected populations to develop a well-mixed gene pool). Thus, in the Ponderosae, every population should be seen as in some ways unique, separable from other populations on molecular genetic grounds but often, also, on morphological, ecological, or other grounds. Therefore a really accurate understanding of evolutionary relationships within this species-rich section of Pinus requires detailed examination of a rapidly evolving literature of molecular and other studies. I will attempt to review this material, but bear in mind that this is only the tip of the iceberg, and there is a huge and rapidly expanding amount of information on this topic.
Returning to Pinus ponderosa itself, there are in this treatment three subspecies. This approach is consistent with the nuclear genetic work of Willyard et al. (2021a), although mitochondrial studies indicate complexities within each subspecies, with mitochondrial haplotypes varying significantly within the range of the species (Potter et al. 2013, Willyard et al. 2021b). These subspecies are:
The type specimen of P. ponderosa was discovered by David Douglas in 1826 near what is now Spokane, Washington (Little 1980) and described by Lawson in 1836. Callaham (2013) describes a rationale for disregarding the type and designates a neotype collected on the Kettle River, Washington, certainly a very typical location for this species, and a spot that Douglas likely visited.
Molecular clock analyses (Lascoux et al. 2004, cited by Potter et al. 2013) suggest that the separation between P. ponderosa and P. scopulorum occurred more than 250,000 years ago, and the division may have persisted since the onset of glacial/interglacial cycles at the Pliocene/Pleistocene boundary, or even earlier, with separation happening when tectonism created the Basin and Range structure of central Nevada in an area that formerly was montane plateau (see discussion by Potter et al. 2013).
Description
Evergreen monoecious trees to 50(-83) m tall and 120(-265) cm DBH. Stem usually straight and robust; branches thick and persistent; usually one whorl per year; usually 3-6 in a whorl; branches inserted at an angle of 51°-56° from apex of the stem of young trees. Bark on mature trees vertically fissured, dark brown or dark gray to black; on old trees becoming pale yellow to yellowish tan, thick, with broad plates separated by shallow fissures. Xylem resin odor (discernible in bark crevices warmed by the sun), usually like commercial turpentine, sometimes lemon rind, never vanilla or pineapple. Needles finely serrate, sharp-tipped, finely lined with stomata on all faces, 13-25 cm long, yellow-green to blue-green, paler and shorter at higher elevations and under xeric conditions; generally not glaucous, but becoming glaucous at higher elevations and with the onset of winter; flexible and in fascicles of three; remaining green on stem an average of 1.4-7.6 years, remaining green longer at higher elevations; enclosed at the base by sheaths of papery bracts that are, in the first year, 10-15% of needle length but that weather away to leave a thick, dark brown to black sheath 4-8 mm long on the oldest fascicles. Vegetative buds covered with loosely appressed ovate acute scales that are conspicuously fringed with short brown hairlike projections; scales dark reddish brown to light gray-brown, dotted with resin during summer, sometimes covered with resin during winter. Vegetative shoots green or brownish green and sticky, elongating once each year. Pollen cones borne in clusters of 5-30, dark red or reddish-brown to reddish purple, long-cylindrical, 20-90 mm long. Seed cones in first year 15-25 mm long, on stalks half as long; red on emergence, turning greenish brown to drab gray, with short, appressed, mucronate scales; prickles usually erect; when mature, symmetrical and straight to slightly curved, 90-100 mm long, long-ovoid to globose; color during second summer from apple green to reddish brown to dark purple; subsessile; deciduous during the first winter after ripening, with a very short stalk and a few basal scales usually persisting temporarily on branch after the cone falls. Seed cone scales 134-159, slightly concave, usually rounded at apex; apophysis yellow to brown, lustrous, slightly elevated along a transverse keel; umbo salient and forming base of a stout, persistent prickle usually directed outward, but occasionally slightly reflexed; adaxial surface slightly concave and dull tawny to red brown; abaxial surface dark brown, purplish, or blackish. Seeds dark brown or speckled; acute-ovoid; width about two-thirds of length, averaging 4-5 x 6-8 mm. Seed wings 3.3-4.3 times length of seed, length 16-25 mm, thin, semitransparent, pale brown, sometimes darkly streaked with brown, widest at or slightly below midpoint, apex rounded (Callaham 2013a).
The following table is useful in distinguishing the 5 taxa commonly called "ponderosa pine" (based on data reported by Callaham 2013a and Kral 1993). In this table, note that although the differences shown are consistent and statistically highly significant, nonetheless a considerable number of cones and leaf fascicles may have to be measured in order to confidently distinguish these taxa. Be careful to select fresh and typical plant material and see the foregoing description regarding differences that may occur at high elevations or on very dry sites. Finally, note that artificial hybrids are possible between all of these taxa and the extent of naturally occurring hybrids is not well known. Some specimens will not be confidently assignable to a taxon.
| Character |
P. ponderosa subsp. benthamiana |
P. ponderosa subsp. ponderosa |
P. ponderosa subsp. washoensis |
P. brachyptera |
P. scopulorum |
| Needle length (cm) |
22.8-27.5 |
24.0-28.4 |
10-17 |
19.1-24.5 |
19.4-22.8 |
| Needles / fascicle |
3.0 |
3.0 |
3.0 |
3.0 |
2.3-2.7 |
| Seed cone length (mm) |
98.9-103.9 |
87.5-89.9 |
70-100 |
72.4-77.4 |
68.5-73.6 |
| Seed cone width / length |
0.77-0.83 |
0.81-0.87 |
<0.67 |
0.84-0.88 |
0.88-0.93 |
| Vegetative shoots |
green or brownish green and viscid |
green or brownish green and viscid |
orange |
reddish brown with blue or purple waxy bloom |
brown or with white to gray waxy bloom |
| Pollen cones |
35-90 mm, dark reddish brown to reddish purple |
20-70 mm, dark red to reddish purple |
10-20 mm, red to purple |
[not stated] |
10-mm, yellow, pink, red to purple |
Distribution and Ecology
Canada: S British Columbia; US: Washington, Oregon, Idaho, Montana, California, Nevada. See the range map shown above. Mostly in the mountains, in pure stands or mixed conifer forests (Little 1980). See also Thompson et al. (1999). Hardy to Zone 4 (cold hardiness limit between -34.3°C and -28.9°C) (Bannister and Neuner 2001, subspecies not specified), but I would expect this to vary significantly according to subspecies and provenance.
Distribution data for the various taxa that have been called "Pinus ponderosa", i.e. P. arizonica, P. brachyptera, P. ponderosa, and P. scopulorum, including subspecies and varieties. Data downloaded on 2021.12.15 from the Consortium of California Herbaria, the Consortium of Midwest Herbaria, and the Consortium of Pacific Northwest Herbaria, limited to geolocated records with coordinate uncertainty less than 10 km. Note that there are probably some misidentifications; for instance, P. scopulorum is unlikely to be found in the North Cascades. Also, there is only one record of P. brachyptera, but genetic studies have found it to be a widespread taxon in Arizona and some neighboring areas; many records attributed to P. ponderosa or P. scopulorum would likely key out to P. brachyptera. Most records include a link to the original herbarium record.
Ponderosa pine presents one of the best examples of the superb adaptation to wildfire that characterizes much of the genus Pinus. Mature ponderosas are not harmed by low intensity fires due to their thick, fire resistant bark, while ponderosa seedlings also have an excellent chance of surviving low-intensity burns because their rootstocks often survive. In the historical period, an enthusiastic program of fire suppression has virtually eliminated these small, frequent fires. As a consequence, shrubs and shade-tolerant conifers have invaded many ponderosa stands while thick accumulations of highly combustible fuels have built up on the forest floor. Now, when a fire does occur, it is likely to be extremely destructive, destroying vast stands of prime forest. Since the mid-1970s, some forest managers have attempted to reintroduce low intensity fire to this ecosystem, but their efforts are often thwarted both by a "Smokey the Bear" mentality ingrained in the public mind, and by the high cost of monitoring prescribed fire in an ecosystem that has accumulated high fuel loads (ironically, there always seems to be plenty of funding to fight the catastrophic fires that result from the absence of fuels control burns).
The same problem can also be viewed from a historical perspective. Early travelers in the West often commented on the vast, parklike stands of ponderosa pine that they encountered. There are tales of stands of giant trees separated by grassy swards, with no other understory vegetation. Many of these stands were so open and level that a carriage could easily be driven through them, and some writers report stands of this character so extensive that they took days to ride across. Such a stand is shown in the photograph at right, taken in the woods near Whitney, Oregon in about 1900. That forest was logged long ago. Comparable forest structure is now being achieved within the range of this species as a result of several decades of forest restoration efforts; the mixed conifer forests in the montane zones of Yosemite, Sequoia and Kings Canyon National Parks provide illustrations. Interestingly, similar historical accounts describe 19th century forests of Pinus palustris in the southeastern U.S., although the trees were not so massive as those seen in the photograph at right. In each of these forests, the giant trees and open understory were a legacy of frequent, low-intensity fire that excluded competitors, discouraged root rot fungi, and maintained an optimum growing environment for the dominant canopy tree species.
The IUCN does not recognize this as a species distinct from P. brachyptera and P. scopulorum and their conservation assessment for that species conflates the three taxa. However, it is still probably accurate to assess P. ponderosa as "least concern" under IUCN criteria due to its extensive area of occupancy and the limited scope of its threats.
Remarkable Specimens
The largest specimens are found in subspecies benthamiana. The tallest specimens are probably also subsp. benthamiana, but occur in an areas where both benthamiana and ponderosa are reported, and have not been conclusively identified as to subspecies. The tallest specimens have all been discovered since about 2010 by Michael Taylor using review of LIDAR data (see photo at right) covering essentially all sites with good potential to have very tall ponderosas (M. Taylor e-mail 2022.04.18). The five tallest known specimens all occur in a fairly small area within Siskiyou National Forest in southwest Oregon, and have heights ranging from 81.33 to 82.97 m, with diameters of 137 to 173 cm. All 50 of the tallest known ponderosas occur in southwest Oregon or extreme northern California, in a broad band on the western slope of the South Cascades and reaching west to the inland portions of the Siskiyous. Historical surveys also found the tallest ponderosas in this region, so the area clearly has conditions conducive to the growth of very tall trees. What those conditions are, has not been demonstrated. I would infer that important factors include closed-canopy forest that drives trees to put on height in order to overtop their neighbors; ample soil moisture during the summer drought; and a low to moderate severity fire regime that maintains ponderosa dominance with minimal mortality. These conditions are likely to change with forecast climate change during the remainder of the 21st century.
The oldest trees are found in subspecies ponderosa, which has attained 907 years (ring count).
Ethnobotany
The Nez Perce indians used to harvest the inner bark to feed their horses in early spring when better forage was still buried deeply beneath the snows. The cambium would be scraped for food (often only in times of famine) by the Klamath, Sanpoil, Spokan, Colville, Okanogan, and Thompson tribal peoples. Nearly everyone ate the seeds, usually roasted, sometimes ground into a flour. The pitch or sap was used by many tribes including the Cheyenne, Flathead, Okanogan, Colville, Paiute, and Thompson for a wide variety of purposes including as a salve or ointment for sores, boils, cuts and earache; to reduce inflammation of the eyes; to treat backache or rheumatism; and to pacify babies by spreading it on their skin. Decoctions of green leaves were put to similar uses. Many tribes used the pitch as a glue or sealer. The Diegueño would make baskets from the needles and the Karok and Maidu would do the same with the roots. The wood was widely used in structures, to make dugout canoes, and for other uses such as cradle boards, ladders, and of course firewood (University of Michigan Ethnobotany Database 2013.09.07).
Currently, this is the most commercially important western pine (Little 1980). Common current uses include structural lumber, lumber for furniture and finish carpentry, firewood, and log cabin construction. It is "generally recognized as the most versatile wood found in abundance in North America" (Western Wood Products Association 2018).
Subspecies benthamiana and ponderosa have both been the subject of many dendrochronology studies, addressing topics as diverse as climate change, ecological succession, ethnobotanical use, and air pollution impacts.
Observations
See the subspecies descriptions.
Remarks
The epithet ponderosa means "large".
Perhaps no other tree is as emblematic of the American west. Within its range, one can seldom travel far without seeing a ponderosa (or, it is often called, "yellow pine"). It has a wide ecological amplitude; Muir (1894) found it "growing bravely upon well-watered moraines, gravelly lake basins, arctic ridges, and torrid lava-beds; planting itself upon the lips of craters, flourishing vigorously even there, and tossing ripe cones among the ashes and cinders of Nature’s hearths." The ponderosa built settlers' cabins and furniture, fueled steam locomotives, timbered mines, and fed sawmills throughout the rise of this country. Even today its management is the first concern of forest management across much of its range, and endless studies have been published on the species' fire ecology, its response to climate change, its vulnerability to diseases, the ecological functions of every life history stage from seedlings to old growth, and countless other attributes of the species' biology. Books, poems, songs, paintings and photographs are dedicated to this species.
Quail, squirrels and many other kinds of wildlife consume the seeds, and nutcrackers and chipmunks cache them, thereby helping to bring forth new pines (Little 1980).
Ponderosa pine (Pinus ponderosa) is the state tree of Montana (Kral 1993).
Citations
Conkle, M. and W. B. Critchfield. 1988. Genetic variation and hybridization of ponderosa pine. Pp. 27-44 in D. Baumgartner and J. Lotan (eds.) Ponderosa pine: the species and its management. Pullman, WA: Washington State University.
Critchfield, W. B. 1966 Crossability and relationships of the California big-cone pines. USDA Forest Service Research Paper NC-6:36–44.
Lascoux, M., A.E. Palme, R. Cheddadi, and R.G. Latta. 2004. Impact of Ice Ages on the genetic structure of trees and shrubs. Philosophical Transactions of the Royal Society of London. B, Biological Sciences 359:197-207.
Lawson, P. 1836. Agriculturist's Manual. Edinburgh (p. 354).
Potter, K.M., V.D. Hipkins, M.F. Mahalovich, and R.E. Means. 2013. Mitochondrial DNA haplotype distribution patterns in Pinus ponderosa (Pinaceae): range-wide evolutionary history and implications for conservation. American Journal of Botany 100(8):1-18.
Silba, J. 2009. Journal of the International Conifer Preservation Society 16(1): 30.
Western Wood Products Association. 2018. Ponderosa pine. www.wwpa.org/western-lumber/species/ponderosa-pine, accessed 2018.10.03.
Willyard, Ann, David S. Gernandt, Blake Cooper, Connor Douglas, Kristen Finch, Hassan Karemera, Erik Lindberg, Stephen K. Langer, Julia Lefler, Paula Marquardt, Dakota L. Pouncey, and Frank Telewski. 2021a. Phylogenomics in the hard pines (Pinus subsection Ponderosae; Pinaceae) confirms paraphyly in Pinus ponderosa, and places Pinus jeffreyi with the California big cone pines. Systematic Botany 46:538–561.
Willyard, A., Gernandt, D.S., López-Reyes, A., and Potter, K.M. 2021b. Mitochondrial phylogeography of the ponderosa pines: widespread gene capture, interspecific sharing, and two unique lineages. Tree Genetics & Genomes 17(6):47. doi:10.1007/s11295-021-01529-4.
Wood Magazine. 2018. Ponderosa pine. www.woodmagazine.com/materials-guide/lumber/wood-species-3/ponderosa-pine, accessed 2018.10.03.
M.P. Frankis contributed much to the development of this page in 1998.12.
See also
Allen, Craig D. 1995. A Ponderosa Pine Natural Area Reveals Its Secrets. In Status and Trends of the Nation's Biological Resources. USGS electronic publication. http://biology.usgs.gov/s+t/SNT/noframe/sw153.htm, accessed 2002.09.03.
Burns & Honkala 1990.
Callaham, R. Z. 2013a. Pinus ponderosa: a taxonomic review with five subspecies in the United States. Research Paper PSW-RP-264, USDA Forest Service, Pacific Southwest Research Station, Albany, CA.
Callaham, R. Z. 2013b. Pinus ponderosa: Geographic races and subspecies based on morphological variation. Research Paper PSW-RP-265, USDA Forest Service, Pacific Southwest Research Station, Albany, CA.
Douglass, Andrew Ellicott. 1919. Climatic Cycles and Tree Growth. Washington, DC: Carnegie Institute of Washington. 127 pp.
Elwes and Henry 1906-1913 at the Biodiversity Heritage Library. This series of volumes, privately printed, provides some of the most engaging descriptions of conifers ever published. Although they only treat species cultivated in the U.K. and Ireland, and the taxonomy is a bit dated, still these accounts are thorough, treating such topics as species description, range, varieties, exceptionally old or tall specimens, remarkable trees, and cultivation. Despite being over a century old, they are generally accurate, and are illustrated with some remarkable photographs and lithographs.
Epperson, B.K., Telewski, F.W., Plovanich-Jones, A.E., and Grimes, J.E. 2001. Clinal differentiation and putative hybridization in a contact zone of Pinus ponderosa and P. arizonica (Pinaceae). American Journal of Botany 88(6): 1052–1057.
FEIS database.
Fiedler, Carl. E. and Stephen F. Arno. 2015. Ponderosa: People, Fire, and the West's Most Iconic Tree. Mountain Press Publishing Company, 260 pages.
Geada López, G., Kamiya, K., and Harada, K. 2002. Phylogenetic relationships of diploxylon pines (Subgenus Pinus) based on plastid sequence data. International Journal of Plant Sciences 163(5): 737–747. doi:10.1086/342213.
Gernandt, D.S., Geada López, G., García, S.O., and Liston, A. 2005. Phylogeny and Classification of Pinus. Taxon 54(1): 29. doi:10.2307/25065300.
Gernandt, D.S., Hernández-León, S., Salgado-Hernández, E., and Rosa, J.A.P. de la. 2009. Phylogenetic relationships of Pinus subsection Ponderosae inferred from rapidly evolving cpDNA regions. Systematic Botany 34(3): 481–491.
Haller, R., and Vivrette, N. 2011. Ponderosa Pine Revisited. Aliso 29(1): 53–57. doi:10.5642/aliso.20112901.07.
Hernández-León, S., Gernandt, D.S., Pérez de la Rosa, J.A., and Jardón-Barbolla, L. 2013. Phylogenetic Relationships and Species Delimitation in Pinus Section Trifoliae Inferrred from Plastid DNA. PLoS ONE 8(7): e70501. doi:10.1371/journal.pone.0070501.
Lauria (1991, 1996a, 1996b).
Murphy, Alexandra. 1994. Graced by Pines: The Ponderosa Pine in the American West. Mountain Press Publishing Company, 119 pages.
Potter, K.M., Hipkins, V.D., Mahalovich, M.F., and Means, R.E. 2015. Nuclear genetic variation across the range of ponderosa pine (Pinus ponderosa): Phylogeographic, taxonomic and conservation implications. Tree Genetics & Genomes 11(3): 38. doi:10.1007/s11295-015-0865-y.
Willyard, A., Cronn, R., and Liston, A. 2009. Reticulate evolution and incomplete lineage sorting among the ponderosa pines. Molecular Phylogenetics and Evolution 52(2): 498–511. doi:10.1016/j.ympev.2009.02.011.
Willyard, A., Gernandt, D.S., Potter, K., Hipkins, V., Marquardt, P., Mahalovich, M.F., Langer, S.K., Telewski, F.W., Cooper, B., Douglas, C., Finch, K., Karemera, H.H., Lefler, J., Lea, P., and Wofford, A. 2017. Pinus ponderosa: A checkered past obscured four species. American Journal of Botany 104(1): 161–181. doi:10.3732/ajb.1600336.