P. caribaea var. bahamensis in habitat, Lucayan National Park, Bahamas [Vladimir Dinets, 2013.04.21].

Trees in cultivation in Caribbean mountains (Carrington, no date).

Foliage on a tree at Quail Botanical Garden, CA [C.J. Earle, 2004.04.06].

Cone about 8 cm long on a tree at Quail Botanical Garden, CA [C.J. Earle, 2004.04.06].

Bark of a 30 cm diameter tree at Quail Botanical Garden, CA [C.J. Earle, 2004.04.06].
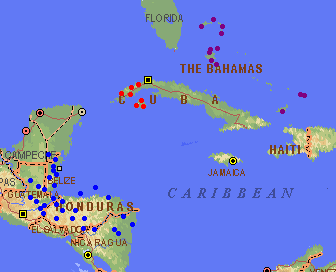
Range of Pinus caribaea (Farjon and Styles 1997). Var. caribaea in red, var. bahamensis in purple, and var. hondurensis in blue. Basemap from Expedia Maps.

P. caribaea var. caribaea planted outside its natural range at relatively high altitudes (900 m foreground to 1200 m background) in the Sierra Maestra, Cuba [Burkhard Witt, 2009].

P. caribaea var. caribaea plantation on good soil within its natural range, province of Pinar del Rio, Cuba [Burkhard Witt, 2009].

Natural regeneration of P. caribaea var. caribaea in a native stand, province Pinar del Rio, Cuba [Burkhard Witt, 2009].

Conservation Status

Pinus caribaea
Morelet 1851
Common names
Pino macho [Spanish]; Caribbean pine; カリビアマツ [Japanese]. Var. bahamensis is called Bahamas pine and var. hondurensis is pino de Honduras.
Taxonomic notes
Occurs in Pinus section Trifoliae, subsection Australes, as do three other major Caribbean pines: P. cubensis, P. occidentalis, and P. elliottii. There are three varieties (Farjon and Styles 1997):
- Pinus caribaea var. caribaea (syn. P. recurvata Rowlee 1903; P. cubensis Grisebach var. anomala Rowlee 1903);
- Pinus caribaea var. bahamensis (Griseb.) W.H. Barrett et Golfari 1851 (syn. P. bahamensis Grisebach 1864);
- Pinus caribaea var. hondurensis (Sénécl.) W.H. Barrett et Golfari 1962 (syn. Pinus hondurensis Sénéclauze 1868; Pinus hondurensis Loock 1950 non Sénéclauze).
Of these, var. hondurensis apparently arose first, with var. caribaea and var. bahamensis evolving somewhat later (Dvorak et al. 2000).
Description
Trees 20-35(-45) m tall and 50-100 cm dbh, usually with a single, round, slender, erect trunk. First-order branches slender and ascending, later ascending to drooping, forming an irregular crown with a crown depth 25-33% of tree height. Inner bark red-brown, bark rough, scaly, fissuring into irregularly square plates. Foliar units in dense, irregularly spaced tufts. Needles in fascicles of (2-)3(-4), 15-26 cm × 1.4-1.8 mm, slightly twisted, rigid, serrulate, acute-pungent, light to dark green, with stomata on all faces; persisting 3 years. Fascicle sheaths persistent, 15-20 mm long at first but weathering shorter. Pollen cones clustered at base of new shoots (often several nodes in one season), 5-6 × 20-30 cm, pink or yellow. Seed cones subterminal, often produced at several nodes in one season, usually in pairs or whorls of 3-5 on 2-2.5 cm long peduncles, maturing in 2 years and opening soon after, ovoid-conical, fairly symmetrical, 5-12 × 4-6 cm when open, with 120-200 seed scales; umbo with a minute, persistent prickle. Seeds variable from light gray-brown to blackish, 5-7 × 2.5-3.5 mm, with a 10-20 mm seed wing. Cotyledons 5-8, (12-)20-35 mm long. Seedlings initially bear primary foliage, eventually developing mature foliage (Farjon and Styles 1997). See García Esteban et al. (2004) for a detailed characterization of the wood anatomy.
Apart from the usual foliage and cone characters, the most noteworthy distinguishing characteristics for this species are the long fascicle sheaths and the multinodal shoots, the latter being an effective adaptation to the tropical climate: when it rains, the tree grows, and when the rains stop, so does the growth.
Farjon and Styles (1997) provide the following key to the varieties of this species:
| | | |
| 1. | Needles in fascicles of (2-)3, fascicle sheaths at maturity reduced in length from 15-20 mm to 10 mm. | var. bahamensis |
| 1. | Needles in fascicles of 3(-4), fascicle sheaths persistently 15-20 mm long. | 2. |
| 2. | Seedlings with green, ascending primary leaves, in a few years replaced by secondary leaves. | var. caribaea |
| 2. | Seedling with glaucous, spreading primary leaves, development of secondary leaves delayed. | var. hondurensis |
| | | |
Distribution and Ecology
Caribbean: Bahamas and Turks-Caicos Islands (var. bahamensis); W Cuba: Pinar del Rio and Isla de la Juventud (var. caribaea); Mexico: S Quintana Roo; N Guatemala; Belize; Honduras incl. Islas de la Bahía; El Salvador; Nicaragua (var. hondurensis) (Farjon and Styles 1997, Dvorak et al. 2000).
Zone 10 (cold hardiness limit between -1°C and +4.4°C) (Bannister and Neuner 2001).
Distribution data from USGS (1999). Var. bahamensis shown in green, var. caribaea in blue, and var. hondurensis in red.
Var. caribaea occurs naturally in westernmost Cuba, in Pinal del Rio and on the Isla de la Juventud (also called Isla de Pinos), which is the type locality. Here it forms pure stands of forest or woodland in a fire climax. The soils are well-drained and acidic, and the elevation is 1-400(-700) m. The trees can grow the year around, but due to the long dry periods, most growth occurs in response to rainfall events (1000-1800 mm/year precipitation); thus the nodal growth described above, and as a consequence, this species does not produce annual rings, instead producing an average of 3-4 rings per year in response to rainfall events. Understory vegetation consists of grasses and scattered shrubs (Farjon and Styles 1997 and other sources).
Var. caribaea has conservation status "vulnerable" (IUCN Red List 2009.10.13). However, Burkhard Witt (email 2009.10.12) disagrees with the IUCN determination for var. caribaea, noting that data reporting the species' decline mostly date to the early 1960's or earlier (INIF-FAO 1962). Since about 1959, about 40,000 ha per year have been reforested in Cuba, and in the province of Pinar del Rio this has mostly been with var. caribaea (Figueroa-Sierra 2002). In his visits to Cuba in 2004 and 2008, Witt found that "Pinus caribaea var. caribaea nowadays in its native range in the province Pinar del Rio is the most common forest tree... Even in the rest of the country Cuba (outside its natural range), in the mountains (e.g. Sierra Maestra, Sierra de Nipe, Sierra del Escambray) plantations of Pinus caribaea var. caribaea are common. In contrast, I see the massive planting of Pinus caribaea var. caribaea as a threat for the species Pinus tropicalis" (Burkhard Witt email 2009.10.12). Most of the stands of Pinus tropicalis were converted to productive Pinus caribaea var. caribaea plantations in the last decades (Figueroa-Sierra 2002). See also Thompson et al. (1999).
Var. bahamensis occurs in the Bahamas on Grand Bahama, Great Abaco, New Providence, North and South Andros Islands, and Great Inagua Island; it also occurs in the Turks-Caicos Islands. It is found on alkaline soils, usually in areas of eroded beach rock and on old beaches, where it forms pure, open, fire-climax stands at altitudes of 1-10 m. Precipitation of 1000-1500 mm annually often comes in the form of hurricanes. Understory vegetation is dominated by Sabal palmetto (Farjon and Styles 1997). See also this description of the Bahamian Pineyards (accessed 2009.10.13).
Var. hondurensis is the most widely distributed variety, occurring widely in Belize and on the Atlantic side of Guatemala, Honduras and Nicaragua; also in Mexico: S Quintana Roo; and at the divide on the El Salvador side of the border with Honduras. This is, in Nicaragua, the southernmost pine in the Americas. It mostly occurs on the coastal plain, bounded below by mangrove swamps and above competition with oaks (Quercus spp.) or other pines, chiefly P. oocarpa or P. tecunumanii, with which it sometimes grows (and sometimes hybridizes). Usually, though, it forms pure stands in a fire-climax forest developed on well-drained, acidic soils at elevations of 1-700 m. Annual precipitation varies greatly, from 660 mm (at Los Limones in interior Honduras) to 4000 mm (at Laguna del Pinar in Nicaragua); there is no frost, but winters comprise the dry season. Forest understory vegetation is variable, predominately including grasses, bracken (Pteridium aquilinum), and Sabal palmetto. (Farjon and Styles 1997). Human influence on the fire regime may have led this species to replace tropical broadleaved forest in many areas (Taylor 1962, cited by Dvorak et al. 2000). This variety is listed as threatened by the Mexican government (NOM-ECOL-059-94), and research by CAMCORE (Dvorak et al. 2000) indicates that the taxon is vulnerable overall due to threatened loss of diversity that would occur with loss of populations at "Guanaja Island and Limón in northern Honduras, the interior dry site provenances in central Honduras extending into El Salvador, the extreme southern populations at Karawala, Pine Ridge Point, and Laguna del Pinar, Nicaragua, and coastal Belize provenances."
Remarkable Specimens
Ethnobotany
This is an important timber tree and is widely exploited; athough the species is not thereby threatened, this history has likely reduced genetic diversity within the taxon, particularly with regard to var. hondurensis (Farjon and Styles 1997).
Observations
For var. bahamensis, Abaco National Park is a good place to see it. This pine forest contains the primary breeding habitat of the endemic Bahama parrot. It can also be seen at Lucayan National Park on Grand Bahama Island.
Remarks
In Belize, P. caribaea var. hondurensis is a host to the dwarf mistletoe Arceuthobium hawksworthii (Hawksworth and Wiens 1996).
Citations
Carrington, Sean. [no date]. The Conifers. http://scitec.uwichill.edu.bb/bcs/bl14apl/gym3.htm (accessed 2004.12.08).
Dvorak, W.S., E.A. Gutiérrez, G.R. Hodge, J.L. Romero, J. Stock and O. Rivas. 2000. Pinus caribaea var. hondurensis. Pp. 12-33 in Conservation & Testing of Tropical & Subtropical Forest Tree Species by the CAMCORE Cooperative. Raleigh, NC: College of Natural Resources, NCSU.
Figueroa Sierra, C. F. 2002. Ecología y conservación de Pinus tropicalis en bosques naturales de las alturas de pizarras. Dissertation, University Pinar del Río (Cuba) and University Alicante (Spain), 124 p.
INIF-FAO. 1962. Seminar and study tour of Latin American conifers. Instituto Nacional de Investigaciones Forestales-FAO. México, D.F. pp. 194-209.
Taylor, B. W. 1962. The status and development of the Nicaraguan pine savannas. Caribbean Forester 23(1):21-25.
See also
Ecosystems of the Bahamas, accessed 2009.10.12.
The species account at Threatened Conifers of the World.
Cordero, J. and D.H. Boshier (eds.). 2004. Pinus caribaea Morelet. Pp. 761-766 in Árboles de Centroamérica. UK: Oxford Forestry Institute (OFI) and Costa Rica: Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). herbaria.plants.ox.ac.uk/adc/downloads/capitulos_especies_y_anexos/pinus_caribaea.pdf [in Spanish], accessed 2011.02.25.
Myers, R., D. Wade, and C. Bergh. 2004. Fire management assessment of the Caribbean pine (Pinus caribea) forest ecosystems on Andros and Abaco Islands, Bahamas. Arlington, VA: The Nature Conservancy. Available: http://www.treesearch.fs.fed.us/pubs/21640, accessed 2009.10.12.
Nelson, C. D., Nance, W. L. and Wagner, D. B. 1994. Chloroplast DNA variation among and within taxonomic varieties of Pinus caribaea and Pinus elliottii. Canadian Journal of Forest Research 24:424-426.
Nieto, V. M. and J. Rodriguez. 2003. Pinus caribaea. Species description in the Tropical Tree Seed Manual. Available http://www.rngr.net/Publications/ttsm/Folder.2003-07-11.4726, accessed 2007.08.31.
Nikles, D. G. 1966. Comparative variability and relationship of Caribbean pine (Pinus caribaea Mor.) and slash pine (Pinus elliottii Engelm.). Ph.D. dissertation, North Carolina State University. 201 pp.