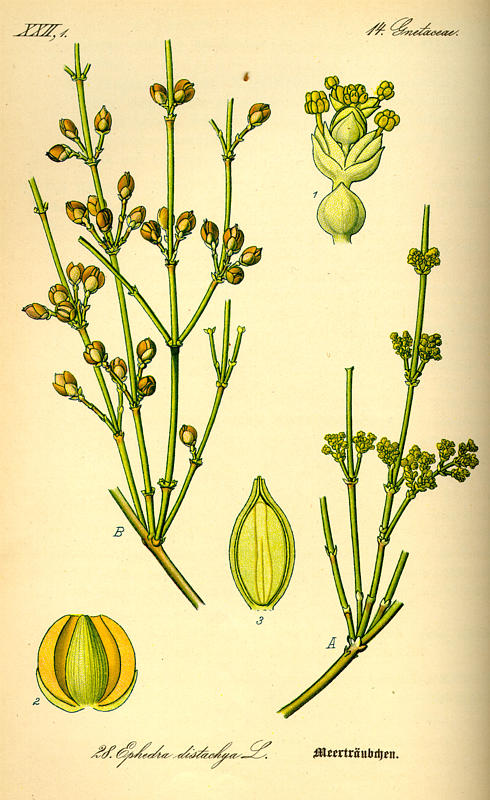
Ephedra distachya [Otto Wilhelm Thomé] (Stüber 1999).
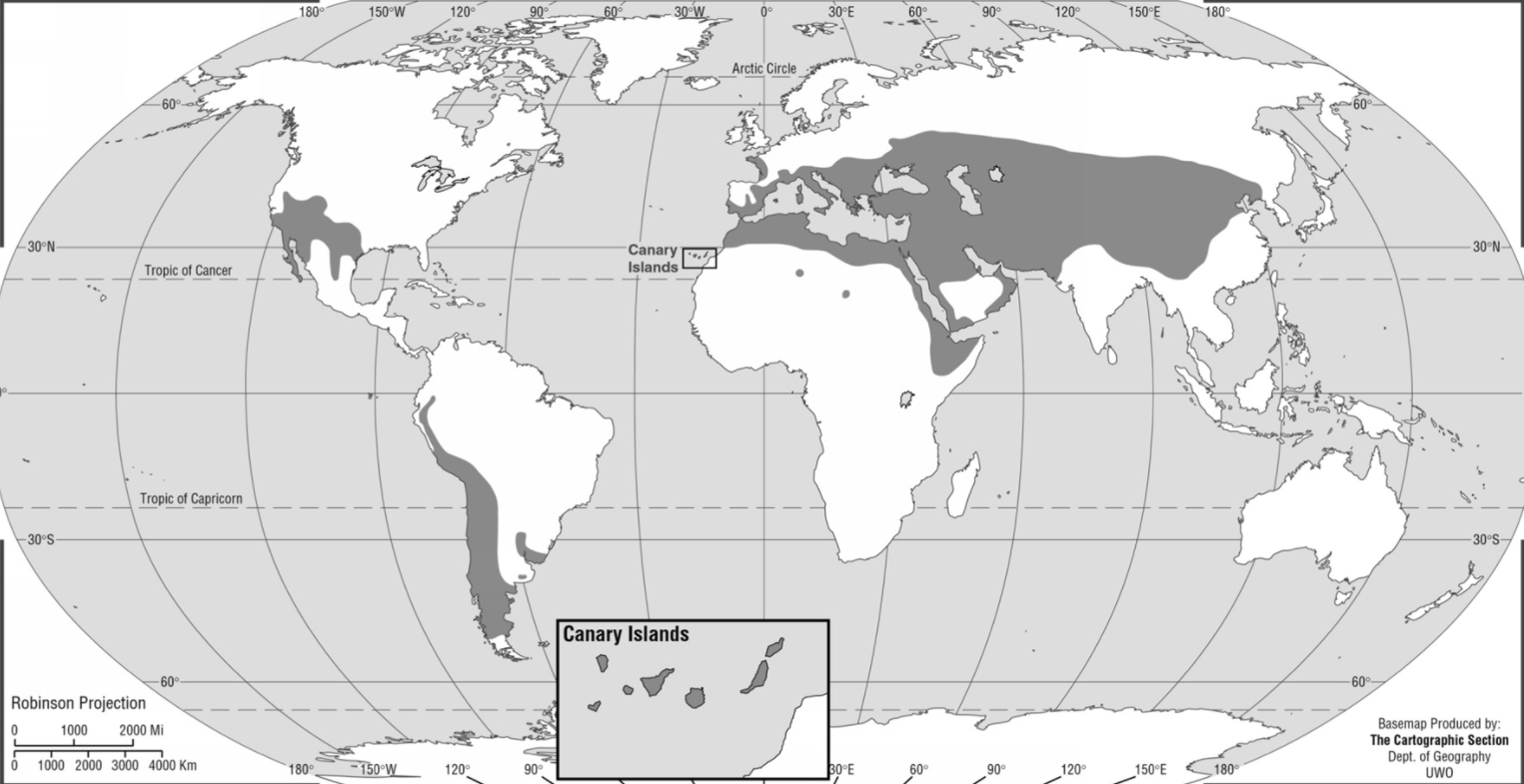
Distribution of Ephedra (Caveney et. al. 2001).
![]()
Ephedra distachya [Otto Wilhelm Thomé] (Stüber 1999).
Distribution of Ephedra (Caveney et. al. 2001).
![]()
Ephedra
Mormon-tea, joint-fir, cañatilla, popotillo, tepopote (Stevenson 1993), 麻黄属 ma huang shu [Chinese] (Fu et al. 1999).
Ephedra is the sole genus in Ephedraceae Dumortier 1829, which some authors designate as the sole family in Order Ephedrales Dumortier 1829 (or, as I prefer, one of 3 families in Order Gnetales Luersson). Its position among the gnetophytes is almost universally accepted, where it is part of a monophyletic clade sister to the genera Gnetum and Welwitschia (Ickert-Bond and Renner 2016). Phylogenetics in the genus are highly correlated with geography, with distinct groups in the Americas, Asia, and the Europe-Mediterreanean region (Huang et al. 2005, Rydin and Korall 2009).
Ickert-Bond and Renner (2016) summarize phylogenetic relationships within Ephedra as follows: "Molecular phylogenies of Ephedra by now have included 56 species. In combination, they show that the six Mediterranean species form a grade at the base of the phylogeny, while the remaining 50 species form a well-supported clade. All 22 New World species form a clade that is nested within a paraphyletic Old World grade. Within the New World clade, the North American E. pedunculata is the earliest diverging species and sister to two clades, one with ten South American species and one with eleven North American ones." They note that a molecular clock study indicates divergence of the Old and New World taxa in the Oligocene.
The family has been around at least since the early Cretaceous, at which time it contained at least one genus (Ephedrispermum) in addition to Ephedra (Rydin et al. 2006). The surviving species seem to be a relict group that happens to have done well during the climate changes, principally the development of very extensive semiarid habitats, that have swept the globe since the Oligocene.
Unlike other gymnosperms, the chloroplast genome in Ephedra is inherited maternally (Mogensen, 1996); as a result chloroplast data relate gene spread via seed dispersal, rather than pollen dispersal.
With 51 species evaluated, polyploidy has been detected in over three-quarters of Ephedra species. This is the highest incidence of polyploidy observed in gymnosperms. There are 13 diploid and 24 polyploid taxa (4X to 8X), with 18 taxa showing intraspecific incidence of polyploidy. The polyploids have variously arisen through both interspecific hybridization and autopolyploidy, and the frequency of asexual reproduction (through rhizomes) has been cited as a factor influencing the relative abundance of polyploidy in the genus (Ohri 2021).
POWO regards the following 71 species as valid. This is probably an excessive number; it includes a substantial number of recently-described species that may eventually reduced to synonymy with existing taxa; Ickert-Bond and Renner (2016), for instance, recognize 54 species, about equally divided between Old World and New World taxa. The underlined links shown below are taxa of doubtful validity. Besides these, there are 3 described nothospecies, all quite rare:
List of taxa:
Perennial, dioecious (rarely monoecious), erect, procumbent or climbing shrubs or vines; occasionally epiphytic. Bark gray to reddish brown, cracked and fissured, often fibrous. Branching varies widely and is diagnostic; most often it is broom-like with nearly parallel and fastigiate to ascending green stems, but in many species the twigs are densely interfingering and in some they are pliable and pendent. The twigs are photosynthetic, yellowish green to olive-green when young, round, finely longitudinally grooved, and jointed, with the internodes usually 1-10 cm long. In cross-section the twigs reveal stomata and intercellular spaces. Roots are generally fibrous; some species reproduce via runners. Leaves are mostly not photosynthetic, and are opposite or in whorls of 3 at the nodes, simple, scalelike, connate at the base to form a sheath, generally early-deciduous; resin canals are absent. Cotyledons 2. The reproductive structures anatomically resemble conifer cones, in that they consist of a number of bracts whorled or decussate around a central axis, with pollen exposed in the pollen cones and ovules open to the air in seed cones. These structures are also often called male and female strobili, or microsporangiate and megasporangiate strobili, which are general terms that can be applied to the reproductive structures of all gymnosperms (except female Ginkgo plants). The ovoid pollen cones are in whorls of 1-12 at the nodes. Each cone is composed of 2-8 sets of opposite or whorled membranous bracts, the proximal bracts empty, the distal bracts basally fused and fertile. When the cone is fertile, 2-15 yellow sporangiophores, analogous to the stamens of flowering plants, each with 2-3 terminal microsporangia, protrude from between the bracts and bear pollen. The pollen is ellipsoidal, with 6-12 longitudinal furrows, not winged, average diameter 27–58 µm. Most Ephedra species are wind-pollinated, but two species basal to the Ephedra clade (E. aphylla and E. foeminea) are insect pollinated (by flies and/or moths), and the other two families of the Gnetales are exclusively insect-pollinated, indicating that wind pollination developed independently in Ephedra. There are three types of seed cones. The first type (Sect. Alatae) bears free, dry, winged, membranous bracts. Second, Sect. Asarca has free, dry, coriaceous bracts. Third, Sect. Ephedra has thickened, colorful, and fleshy bracts. Sections Alatae and Asarca are nested within Sect. Ephedra, suggesting that the fleshy cone is basal in Ephedra. The seed cones are also structures of paired bracts formed in whorls of 1-10 at the nodes; when fertile, seed cones are smaller than the pollen cones. Each cone is sessile or on a peduncle, composed of 2-10 sets of overlapping, opposite or whorled bracts, the proximal bracts empty, the most distal bracts subtending 1 axillary cone composed of a pair of fused bracteoles enclosing a single-integumented ovule with the integument projecting as a tube (the micropylar tube) through which the ovule is fertilized. This tube projects from the bracteole envelope, which forms a leathery "seed coat" that is shed with the seed. If fertilized, typically only one ovule develops a seed. The seeds are dispersed by wind, birds, or rodents, with different dispersal mechanisms predominating in different species. There are 1-2(-3) seeds per cone, yellow to dark brown, smooth or furrowed. The wood is ring porous, lacking resin ducts, with wide multiseriate rays and vessels in older stems; the vessels are most abundant and largest in lianoid species, but are nearly absent in alpine species (Stevenson 1993, Fu et al. 1999, Rydin and Bolinder 2015, Ickert-Bond and Renner 2016).
Ephedra is generally dioecious and there have been a number of errors in which type specimen collections included a male of one species and a female of another (e.g., E. aphylla). For proper identification of Ephedra, fruiting and flowering material is essential. Many sterile specimens of various taxa look alike (Ali and Qaiser 1987), though some vegetative characters are quite useful, such as the length of the scale leaves attached at nodes.
Deserts, semideserts, desert steppes, or seasonally dry habitats such as mediterranean-type evergreen or deciduous woodlands and subtropical thorn scrub; in both the Old and New Worlds. Often forms clonal colonies (Ickert-Bond and Renner 2016).
The various species are distributed as follows (note that species numbers are approximate):
Their habitats are all described as dry, rocky and/or sandy. A few species occur in grasslands, and for a few species, habitat is not specified.
One species occurs in Argentina and Chile, from Tierra del Fuego to 42° S.
Two species occur in North Africa, one of which also occurs in SW Asia (Saudi Arabia, Kuwait, Israel) and Cyprus.
Twelve species are in the USA (Arizona, California, Colorado, Nevada, New Mexico, Oklahoma, Oregon, Texas, Utah, Wyoming) of which 5 species also occur in Mexico (Baja California Norte, Chihuahua, Coahuila, Nuevo León, San Luis Potosí, Sonora).
The remaining 21 species are Eurasian, with focal areas in central Asia (18 species) and around the Mediterreanean (4 species, plus the North African ones). These break out according to country as follows:
The New World species occur at elevations ranging from below sea level in Death Valley to about 5000 m for E. rupestris in the Andes of Ecuador. The Old World species are found from sea level to 5300 m elevation (E. gerardiana, the highest elevation occupied by any gymnosperm species). E. intermedia probably has the greatest elevational range of any single gymnosperm species, ranging from 100 to 4600 m elevation across its vast range.
The Ephedraceae are the only widespread group of gymnosperms that are not of conservation concern. The IUCN (2020) has not identified any taxa as critically endangered, endangered, or vulnerable, and only one is listed as "near threatened." This happy situation is probably due to the fact that Ephedra generally occupies arid and semiarid habitats that are poorly suited for agriculture or development, is unpalatable to livestock, and has large areas of occurrence.
Seed dispersal varies between species; some are dispersed by wind, some by small mammals who cache the seeds, and some (primarily ones with colorful fleshy seed cones) by birds, which eat the cones and defecate the seeds (Hollander and Vander Wall 2009).
E. boelckei is one of the few species that can grow as a tree, reportedly up to 4.5 m tall.
I have found no records of specimen age. I once cut the stem of a large dead specimen, likely E. viridis, and found that it appears to form annual rings; there were 43 of them.
Throughout its distribution it is used by native cultures for a variety of medicinal purposes, including cough medicines, an antipyretic, an antisyphilitic, a stimulant for poor circulation, and an antihistamine. These uses are based on the presence of tannins and alkaloids, particularly ephedrines (Stevenson 1993). In recent times, it has achieved widespread popularity as an 'herbal medicine' used in weight-loss preparations and 'energy' preparations (both uses due to its stimulant effects) and cold and allergy medications (due to the presence of the bronchodilator ephedrine) (Herbal Information Center. [no date]).
The old world species contain variable amounts of ephedrine; curiously it is not present in the New World species, and the psychotropic properties ascribed to "Mormon Tea" and its relatives remain unknown. The oldest drug produced from Ephedra is the Chinese ma-huang, which has been used in Chinese medicine for over 5000 years to treat fever, nasal congestion and asthma. Ma-huang is widely sold in the western world, usually with dubious claims as to its curative values (literature review by Caveney et al. 2001). Ephedra has also been used in the synthesis of methamphetamine (Andrews 1995).
Ephedra is not used in dendrochronology, but there is likely some potential, since it produces growth rings, which in most cases are of roughly annual frequency.
See the species accounts.
Named from the "Greek ep-, upon, and hédra, seat or sitting upon a place; from the ancient name used by Pliny for Equisetum; the stems resemble the jointed stems of Equisetum, the segments of which appear to sit one upon the other" (Stevenson 1993).
"In Ephedra the habit varies greatly even in the same species. Dwarf bushes a few inches high are common (E. distachya, etc.). E. distachya and other species are also represented by tall bushes up to 6 ft. in height, with virgate branches. E. triandra may attain the habit of a small tree with a trunk a foot in diameter. Climbers of the "weaver" type with slender pendent branches are found in E. altissima and E. fragilis: these and other species include forms in which the branches are prostrate.
"Most species, especially in loose soils, spread by means of branching rhizomes which arise from buried nodes of the erect stems; they thus exert a binding effect upon the soils in which they grow. In E. alata, growing in the sand dunes of the Areg formation of Algeria and Tunis, the rhizomes attain a length of several metres and form efficient sand binders. Erect shoots of the normal type arise from the axils of the younger leaf-sheaths. When these shoots are isolated by the breaking or decay of the parent rhizome they differ from seedlings only in the absence of tap-roots" (Pearson 1929).
Ali, S.I. and M. Qaiser (eds.). 1987. Flora of Pakistan. http://www.efloras.org.
Andrews K.M. 1995. Ephedra's role as the precursor in the clandestine manufacture of methamphetamine. Journal of Forensic Science 40: 551-560.
Caveney, Stanley, David A. Charlet, Helmut Freitag, Maria Maier-Stolte and Alvin N. Starratt. 2001. New observations on the secondary chemistry of world Ephedra (Ephedraceae). American Journal of Botany 88:1199-1208. http://www.amjbot.org/cgi/content/full/88/7/1199, accessed 2006.10.18.
Cutler, H. C. 1939. Monograph of the North American species of the genus Ephedra. Annals of the Missouri Botanical Garden 26:373-427. Available: Biodiversity Heritage Library, accessed 2021.12.28.
Dumortier, B.C.J. 1829. Analyse des familles des plantes, avec l'indication des principaux genres qui s'y rattachent Tournay: J. Casterman aîné. P. 11. Available at the Biodiversity Heritage Library.
Herbal Information Center. [no date]. Ephedra (Ephedra sinica). Article posted at the URL http://www.kcweb.com/herb/ephedra.htm, accessed 1999.02.21.
Hollander, J. L. and S. B. Vander Wall. 2009. Dispersal syndromes in North American Ephedra. International Journal of Plant Sciences 170:323-330.
Huang, J., D. E. Giannasi and R. A. Price. 2005. Phylogenetic relationships in Ephedra (Ephedraceae) inferred from chloroplast and nuclear DNA sequences. Molecular Phylogenetics and Evolution 35(1):48-59.
Ickert-Bond, S. M., and S. S. Renner. 2016. The Gnetales: Recent insights on their morphology, reproductive biology, chromosome numbers, biogeography, and divergence times. Journal of Systematics and Evolution 54:1–16.
IUCN. 2020. IUCN Red List version 2020-1: Table 4b: Red List Category summary for all plant classes and families. https://nc.iucnredlist.org/redlist/content/attachment_files/2020_1_RL_Stats_Table_4b.pdf, accessed 2022.09.23.
Mogensen, H. L. 1996. The hows and whys of cytoplasmic inheritance in seed plants. American Journal of Botany 83:383-404.
Ohri, D. 2021. Polyploidy in gymnosperms -- a reappraisal. Silvae Genetica 70:22-38.
Rydin, C., K.R. Pedersen, P.R. Crane and E.M. Friis. 2006. Former diversity of Ephedra (Gnetales): evidence from early Cretaceous seeds from Portugal and North America. Annals of Botany 98(1):123–140.
Rydin, C. and K. Bolinder. 2015. Moonlight pollination in the gymnosperm Ephedra (Gnetales). Biology Letters, doi.org/10.1098/rsbl.2014.0993.
Rydin, C., and P. Korall. 2009. Evolutionary relationships in Ephedra (Gnetales), with implications for seed plant phylogeny. International Journal of Plant Sciences 170(8):1031–1043. doi:10.1086/605116.
Stapf, O. 1889. Die Arten der Gattung Ephedra. Denkschrift der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse 56(2):1–112.
Stöber, Kurt. 1999. Images from Thomé, Flora von Deutschland etc. http://www.mpiz-koeln.mpg.de/~stueber/thome/index.html, accessed 2003.05.01, now defunct.
Benson, L.D. 1943. Revisions of status of southwestern trees and shrubs. American Journal of Botany 30:230-240.
Bolinda, K. 2014. Pollination in Ephedra. Thesis, Stockholms Universitet. Available: https://www.diva-portal.org/smash/get/diva2:707931/FULLTEXT02.pdf, accessed 2021.12.26. Chapters include Pollination mechanisms in the ancient gymnosperm clade Ephedra (Gnetales) and Aerodynamics and pollen ultrastructure in Ephedra (Gnetales).
Carlquist, Sherwin. 1989. Wood and bark anatomy of the New World species of Ephedra. Aliso 12(3):441.
Carlquist, Sherwin. 1992. Wood, bark and pith anatomy of Old World species of Ephedra and summary for the genus. Aliso 13(2):255.
Chaudhri, I.I. 1957. Pakistani Ephedra. Economic Botany 11: 257-262.
Friedman, W. E. 1990. Double fertilization in Ephedra, a nonflowering seed plant: Its bearing on the origin of angiosperms. Science 247:951-954.
Friedman, W. E. 1990. Sexual reproduction in Ephedra nevadensis (Ephedraceae): Further evidence of double fertilization in a nonflowering seed plant. American Journal of Botany 77:1582-1598.
Freitag, H. and M. Maier-Stolte. 1989. The Ephedra-species of P. Forsskal: identity and typification. Taxon 38:545-556.
Freitag, H. and M. Maier-Stolte. 1993. Ephedraceae. In T. G. Tutin (ed.), Flora Europaea, 2nd ed., vol. 1. Cambridge University Press, London, UK.
Freitag, H. and M. Maier-Stolte. 1994. Ephedraceae. In K. Browicz (ed.), Chorology of trees and shrubs in south-west Asia and adjacent regions. Bogucki Publishers, Poznan, Poland.
Hunziker, J.H. 1949. Sinopsis de las species argentinas del genero Ephedra. Lilloa 17: 147-174.
Hunziker, J. H. 1995. The karyotypes of Ephedra ochreata, E. rupestris and E. viridis. Darwiniana 33:369-370.
Ickert-Bond, Stefanie M. 2003. Systematics of New World Ephedra L. (Ephedraceae): Integrating morphological and molecular data. Ph.D. Thesis, Arizona State University, Tempe.
Ickert-Bond, S. M., and M. F. Wojciechowski. 2004. Phylogenetic relationships in Ephedra (Gnetales): evidence from nuclear and chloroplast DNA sequence data. Systematic Botany 29(4):834–849. doi:10.1600/0363644042451143.
Kubitzki, K. 1990. Ephedraceae. In: K. Kubitzki et al., eds. 1990+. The Families and Genera of Vascular Plants. Berlin etc. Vol. 1, pp. 379-382.
Markgraf, F. 1926. Ephedraceae. In: H. G. A. Engler et al., eds. 1924+. Die naturlichen Pflanzenfamilien..., ed. 2. Leipzig and Berlin. Vol. 13, pp. 409-419.
Meyer, C. A. 1846. Versuch einer Monographie der Gattung Ephedra, dirch Abbildung erlautert. Mémoires de l'Académie impériale des sciences de St.-Pétersbourg, Sér. 6, Sci. Math., Seconde Pt. Sci. Nat. 5:35-107. Available: Biodiversity Heritage Library, accessed 2021.12.31.
Mussayev, I. 1978. On geography and phylogeny of some representatives of the genus Ephedra. Bot. Zurn. (Moscow & Leningrad) 63:523-543. [In Russian.]
Parlatore, F. 1868. Gnetaceae. P.352-359 in A. P. and A. L. P. de Candolle (eds.), 1823-1873. Prodromus Systematis Naturalis Regni Vegetabilis.... Paris etc. Vol. 16, part 2.
Price, Robert A. 1996. Systematics of the Gnetales: A Review of Morphological and Molecular Evidence. International Journal of Plant Sciences 157(6), Supplement: Biology and Evolution of the Gnetales (Nov., 1996), pp. S40-S49.
Rydin, C., K. R. Pedersen, and E. M. Friis. 2004. On the evolutionary history of Ephedra: Cretaceous fossils and extant molecules. Proceedings of the National Academy of Sciences. Available: http://www.pnas.org/cgi/content/full/101/47/16571, accessed 2006.03.21).
Tanaka, Toshihiro; Ohba, Koji; Sakai, Eiji. 1995. Comparison of the Constituents of Ephedra Herbs from Various Countries--On Ephedrine Type Alkaloids. Natural medicines 49(4):418.
Yang, Y., B.-Y. Geng, D. L. Dilcher, Z.-D. Chen, and T. A. Lott. 2005. Morphology and affinities of an Early Cretaceous Ephedra (Ephedraceae) from China. American Journal of Botany 92:231-241.
Zhang J.S., Tian S.Z. and Lou C. 1989. Quality evaluation of twelve species of Chinese Ephedra (Ma Huang). Acta Pharmica Sinica 24: 865-871.
Last Modified 2025-01-30