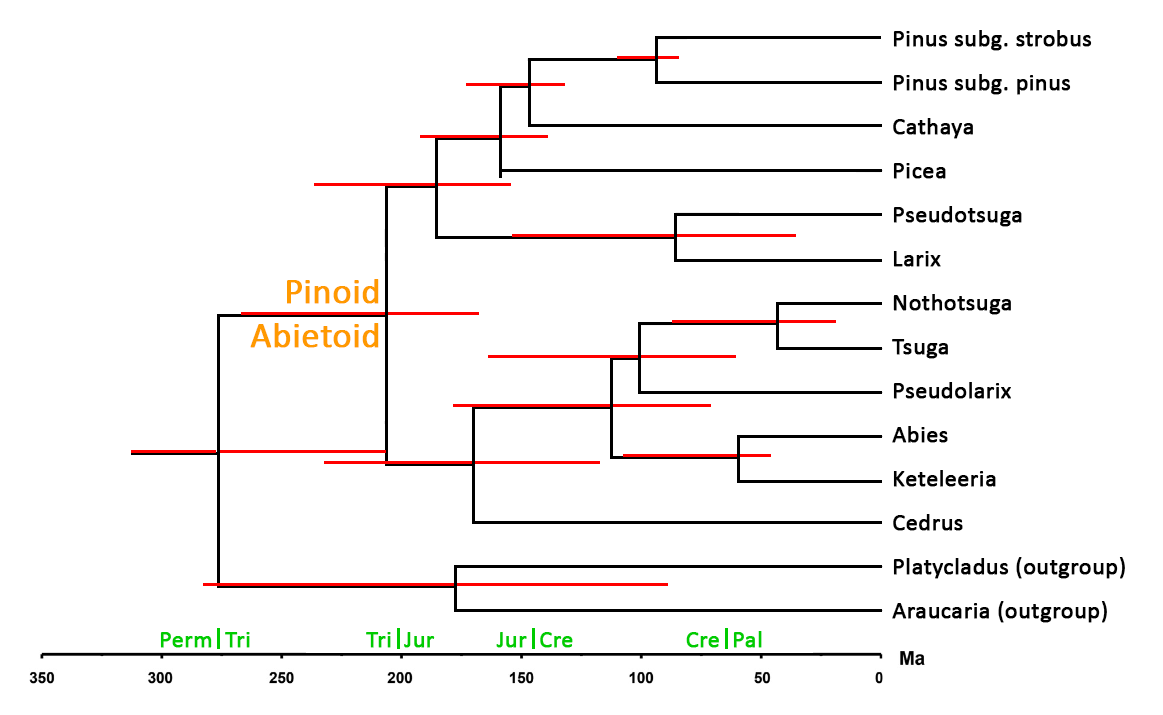
Cladogram of the Pinaceae, with divergence time estimates, based on Figure 5 from Ran et al. (2018b). Red lines indicate 95% posterior probability intervals on divergence times.









![]()
Cladogram of the Pinaceae, with divergence time estimates, based on Figure 5 from Ran et al. (2018b). Red lines indicate 95% posterior probability intervals on divergence times.
![]()
Pinaceae
Pine family, Pinacées [French], Kieferngewächse [German], Pináceas [Spanish], 松科 [Chinese], マツ科 [Japanese].
Sometimes treated as the sole family of Order Pinales Dumortier 1829. Syn: Abietaceae. There are here treated 11 genera and 234 species:
Pinaceae are known in the fossil record since the Cretaceous. The family originally included all conifers but is now restricted to a clearly monophyletic group united by characters seen in the mature seed cones: bract-scale complexes consisting of well-developed scales that are free from the subtending bracts for most of their length, two inverted ovules on the adaxial face of each scale, and a (usually obvious) seed wing developed from the cone scale (Thieret 1993). The family seems to have arisen via a whole-genome-duplication (polyploidy) event that separated it from the remaining conifer lineages (a similar event gave rise to the Cupressaceae-Taxaceae clade, and to the Welwitschiaceae) (Li et al. 2015). Combined molecular and morphological analysis have been used to derive phylogenetics with divergence time estimates. The results, summarized in the cladogram at right, indicate that the family diverged from other conifers about 276 million years ago (ma), with divergence of the pinoid and abietoid clades about 206 ma. There is substantial uncertainty in these early dates due to ambiguities about the relationship to the Gnetales, which may have arisen at about the same time and may be sister to the pinoid clade. Within the pinoid clade, most genera arose during the mid to late Jurassic, though the Larix-Pseudotsuga divergence happened much later in the mid-Cretaceous. In the abietoid clade, divergences happened more sporadically, from mid-Jurassic (Cedrus) to mid-Eocene (Nothotsuga) (Ran et al. 2018a, 2018b). There is large uncertainty in these dates (shown by error bars in the cladogram); amongst the conifers in general, rapid evolution and speciation at generic and lower ranks tends to occur in response to large-scale climate changes, which are often accompanied by mass extinctions. There are also uncertainties due to the absence of a fossil record in the analysis; this is a largely relict group and many taxa, likely including some genera, are missing from the analysis because they are no longer extant.
Cone and seed characters serve to discriminate four subfamilies, as follows (Frankis 1989, Farjon 1990):
Tree: Trees, occasionally shrubs, evergreen (annually deciduous in Larix and Pseudolarix), resinous and aromatic, monoecious.
Bark: Smooth to scaly or furrowed.
Branches: Terminal branches distinct from lateral branches in having radial symmetry. Lateral branches well developed and similar to leading (long) shoots, or reduced to well-defined short (spur) shoots (Larix, Pseudolarix, Cedrus) or dwarf shoots (Pinus); twigs terete, sometimes clothed by persistent primary leaves or leaf bases; longest internodes between leaves less than 1 cm; buds conspicuous. Several genera (e.g. Picea, Pinus, Pseudotsuga, Tsuga) produce epicormic branches, and Pinus produces interfascicular branches.
Roots: Fibrous to woody, commonly comprised of woody structural roots that terminate in nonwoody ectomycorrhizal fine roots that are replaced semiannually to annually.
Leaves: Leaves simple, shed singly (whole fascicles shed in Pinus), alternate and spirally arranged but sometimes twisted at the base so as to appear 1- or 2-ranked, or fascicled, linear to needlelike, sessile to short-petiolate; foliage leaves either borne singly (spirally) on long shoots or in tufts (fascicles) on short shoots; juvenile leaves (when present) borne on long shoots; resin canals present. Cotyledons 2-15(-24).
Cones: Seed cones maturing in 1 season (2-3 seasons in Pinus) and shed soon after, or long-persistent, sometimes serotinous (not opening upon maturity but much later: some Pinus, and Picea mariana), compound, axillary, solitary or grouped. Cones generally lateral, but can be terminal in Picea and Tsuga.
Cone scales: Overlapping, free from subtending included or exserted bracts for most of length, spirally arranged, strongly flattened, at maturity relatively thin to strongly thickened and woody (in Pinus), with 2 inverted, adaxial ovules.
Pollen cones: Matured and shed annually, solitary or clustered, axillary, ovoid to ellipsoid or cylindric; sporophylls overlapping, bearing 2 abaxial microsporangia (pollen sacs); pollen spheric, 2-winged, less commonly with wings reduced to frill (in Tsuga sect. Tsuga), or not winged (in Larix and Pseudotsuga).
Seeds: Two per scale, elongate terminal wing partially decurrent on seed body (wing short or rudimentary in some species of Pinus); aril lacking.
The Northern Hemisphere: south to the West Indies, Central America, Japan, China, Indonesia (one species, Pinus merkusii, crosses the equator in Sumatra), the Himalaya, and North Africa. The family is dominant in the vegetation of large regions including (in North America) forests of the boreal and Pacific regions, the western mountains, and the southeastern coastal plain. Various species, most commonly Pinus radiata, have been widely introduced for timber production in sub-Saharan Africa, South America, New Zealand and Australia.
Worldwide distribution of Pinaceae, generalized to 1x1 degree cells; data from the BRAHMS database at Conifers of the World, accessed 2021.11.11. Note that distribution in boreal regions, especially Siberia, is underrepresented due to limited sampling. Also note that if multiple Pinaceae genera are present in a grid cell, only the least-widespread genus is shown here.
The Pinaceae are, in general, very much of conservation concern. The IUCN (2020) has identified 9 taxa as critically endangered, 25 as endangered, and 24 as vulnerable, thus 26% of all species in the family are at risk. Principal factors of decline, which typically affect all taxa regardless of their conservation listing status, include habitat conversion through agriculture or development, timber harvest, grazing, and climate change.
Big trees are preferably recorded by their wood volume. Calculating wood volume is complex, so dbh is more commonly used. The following table summarizes the largest known individuals in each genus, except, that I have no size data for Cathaya, Keteleeria, or Nothotsuga. Note that Pinus features a tie between the largest species of subgenus Pinus and the largest of subgenus Strobus.
| Genus | Species | Wood volume | DBH | Height |
| Abies | Abies procera | 126 m3 | 253 cm | 82.9 m |
| Cedrus | Cedrus deodara | no data | 460 cm | 45 m |
| Larix | Larix occidentalis | 83 m3 | 221 cm | 64.9 m |
| Picea | Picea sitchensis | 337 m3 | 539 cm | 100.2 m |
| Pinus subg. Pinus | Pinus ponderosa | 153 m3 | 280 cm | 93.0 m |
| Pinus subg. Strobus | Pinus lambertiana | 153 m3 | 318 cm | 78.9 m |
| Pseudolarix | Pseudolarix amabilis (measured 1854) | no data | 150 cm | 40 m |
| Pseudotsuga | Pseudotsuga menziesii | 349 m3 | 485 cm | 99.5 m |
| Tsuga | Tsuga heterophylla | 87 m3 | 276 cm | 83.3 m |
The Pinaceae include Pinus longaeva, the oldest of all non-clonal conifers. Its sister species, P. aristata and P. balfouriana, are the second- and third-oldest in the Pinaceae. See How Old is That Tree? for further discussion.
Both aboriginal and modern uses of the Pinaceae have focused on, first, the timber; then the resins; then horticultural and ornamental plantings; and then, for a relative minority of species, food and spiritual values. There has also been widespread use in science, such as dendrochronology to date past events and chemical studies to assess pharmacological activity. Economically, members of the Pinaceae provide most of the world's softwood timber. They also provide pulpwood, naval stores (e.g., tar, pitch, turpentine, etc.), essential oils, and other forest products. Some species of Pinus produce edible seeds, commonly known by names such as piñon or pignolias. Many species, including most of the genera, are grown as ornamentals and shelter-belt trees and for revegetation.
The genera most commonly seen in cultivation are Abies, Cedrus, Larix, Picea, Pinus, Pseudotsuga, and Tsuga, each of these genera being represented by numerous cultivars. The remaining genera are mainly botanical garden subjects (Thieret 1993).
The majority of all dendrochronological research has involved species of the Pinaceae. Very many studies have involved species of Picea and Pinus, fewer but still a great many studies have involved Abies, Larix, Pseudotsuga and Tsuga, and rather few studies have involved the smaller genera.
See the generic and species descriptions.
The Pinaceae are by far the most economically and ecologically important gymnosperm family. Without the Pinaceae, the gymnosperms are little more than a botanical curiosity, their dominance largely confined to semiarid regions with their abundant Juniperus. With the Pinaceae, the gymnosperms continue to dominate many of the most widespread vegetation types on earth, including most boreal forests, most temperate and boreal mountain forests, and vast expanses of North American, African, European and Asian semiarid woodlands.
Dumortier, B.C.J. 1829. Analyse des familles des plantes, avec l'indication des principaux genres qui s'y rattachent Tournay: J. Casterman aîné. P. 11. Available at the Biodiversity Heritage Library.
Farjon, Aljos. 1990. Pinaceae: drawings and descriptions of the genera Abies, Cedrus, Pseudolarix, Keteleeria, Nothotsuga, Tsuga, Cathaya, Pseudotsuga, Larix and Picea. Königstein: Koeltz Scientific Books.
Frankis, M. P. 1989. Generic inter-relationships in Pinaceae. Notes Roy. Bot. Gard. Edinburgh 45: 527-548.
IUCN. 2020. IUCN Red List version 2020-1: Table 4b: Red List Category summary for all plant classes and families. https://nc.iucnredlist.org/redlist/content/attachment_files/2020_1_RL_Stats_Table_4b.pdf, accessed 2022.09.23.
Li, Zheng, Anthony E. Baniaga, Emily B. Sessa, Moira Scascitelli, Sean W. Graham, Loren H. Rieseberg, and Michael S. Barker. 2015. Early genome duplications in conifers and other seed plants. Science Advances 1(10):e1501084.
Ran, Jin-Hua, Ting-Ting Shen, Ming-Ming Wang, and Xiao-Quan Wang. 2018a. Phylogenomics resolves the deep phylogeny of seed plants and indicates partial convergent or homoplastic evolution between Gnetales and Angiosperms. Proceedings of the Royal Society B: Biological Sciences 285(1881):20181012. doi.org/10.1098/rspb.2018.1012.
Ran, Jin-Hua, Ting-Ting Shen, Hui Wu, Xun Gong, and Xiao-Quan Wang. 2018b. Phylogeny and evolutionary history of Pinaceae updated by transcriptomic analysis. Molecular Phylogenetics and Evolution 129:106–116. doi.org/10.1016/j.ympev.2018.08.011.
Rudolphi, F. 1830. Syst. Orb. Veg., p. 35.
Canadian Forestry Service. 1983. Reproduction of conifers. Forest. Techn. Pub. Canad. Forest. Serv. 31.
This page was co-edited with Michael Frankis, 1999.01.
Last Modified 2025-02-22