Juniperus
Linnaeus 1753, p. 1038
Common names
Juniper, cedar, redcedar; cedro, sabino [Spanish] (Adams 1993).
Taxonomic notes
This is the largest genus in the Cupressaceae in terms of species. In this treatment, there are 73 species:
The genus is characterized by fleshy cones with hard-shelled seeds, which are adaptations to avian seed dispersal. Apart from this, all characters common to all of its species can also be found in other closely allied genera of Cupressaceae, notably Cupressus, Hesperocyparis and Platycladus. Molecular taxonomic methods have been widely applied in the genus, most notably in a very large collection of papers published by Adams and colleagues and mostly published in the open access journal Phytologia, q.v. The first effort to apply a uniform molecular analysis to nearly all species in the genus was published by Mao et al. (2010). Further molecular analyses embracing all or nearly all species in the genus have been performed by Adams (2014) and Leslie et al. (2012), and this group of studies taken together provide a degree of clarity regarding relationships within the genus.
The genus has traditionally been divided into three distinct sections or subgenera, sometimes treated as genera (e.g., by Yang et al. 2023):
- Sect. Juniperus (syn: Juniperus sect. Oxycedrus Spach 1841)
- Sect. Caryocedrus Endlicher 1847 (syn: Arceuthos Antoine 1854)
- Sect. Sabina Spach 1841 (syn: Sabina Miller 1754)
This is one of the few conifer genera that show a high incidence of polyploidy, meaning, in effect, that each plant carries multiple similar but distinct sets of DNA, complicating interpretation of molecular analyses. Farhat et al. (2019) have identified 16 polyploid species, mostly tetraploids and one hexaploid; see their paper for further discussion.
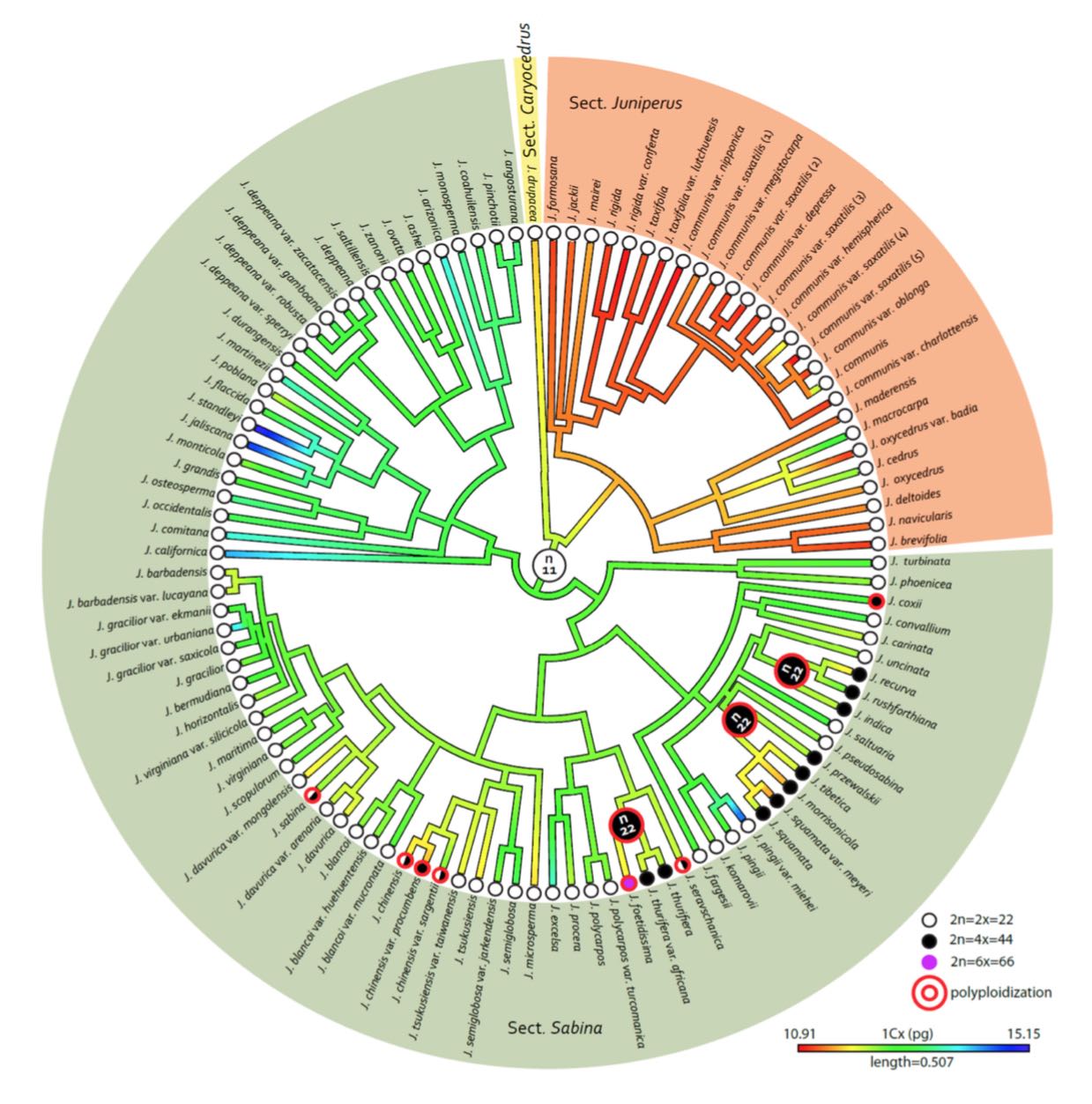
Mao et al. (2010) provided a nearly-comprehensive molecular analysis of the genus; so did Leslie et al. (2012), in an analysis that included estimates of divergence dates; an even more comprehensive analysis (in the sense of including all described taxa) is presented by Adams (2014). Thus we have three more-or-less comprehensive, independently developed molecular phylogenies for the genus; I have used these to generate the cladogram shown here. I also present the circular cladogram presented by Adams (2014), as redrawn by Farhat et al. (2019), in a remarkably informative illustration. In general, all three phylogenies agree in designating seven principal clades within Juniperus, defined as follows, and numbered according to their sequence of divergence.
- Section Caryocedrus; J. drupacea, the only species, differentiated about 20 million years ago.
- Section Juniperus, shown here in magenta, speciation mostly occurring within the past 10 million years.
- Section Sabina, Clade 1 (green) contains species of the Himalaya and Tibetan Plateau.
- Section Sabina, Clade 2 (red) contains the serrate-leaf-margin species of North America, widely recognized as a distinct subgroup.
- Section Sabina, Clade 3 (blue) includes the smooth-leaf-margin American species, which are sister to three earlier-derived subclades containing Asian species.
- Section Sabina, Clade 4 (brown) contains a far-flung mixture of European, Asian and African species.
- Section Sabina, Clade 5 (purple) contains only J. phoenicia and J. turbinata, native to the Mediterranean and the only serrate-leaf-margined Old World species.
In developing this cladogram, I accepted all points on which the three analyses (Adams 2014, Leslie et al. 2012, Mao et al. 2010) agreed. For points on which two analyses agreed, I examined the findings of each of the three studies and based a determination on that. For points an which no two analyses agreed, I conservatively assumed that no phylogenetic relationships can be confidently inferred except to the extent shown here. Thus, for many species, we can say no more than that they belong to a given clade. Also note that Adams (2014) recognizes considerably more species than the other authors; for these I have assumed that Adams' phylogenetic inferences are correct. For further detail on the rationale underlying this cladogram, see remarks in the individual species accounts.
Fossil and molecular clock data presented by Mao et al. (2010) and Leslie et al. (2012) indicate that these major clades differentiated from Cupressus about 50-72 million years ago, during the Paleocene. In the analysis of Mao et al. (2010), subsequent differentiation of the sections in Juniperus and clades in section Sabina occurred at about the time of the Eocene-Oligocene transition, between 30 and 50 million years ago. Disjunctions between Eurasian and North American species arose at three times: 30-43 million years ago during the Eocene-Oligocene transition, when Clade 2 (red) differentiated; 5-17 million years ago within Clade 3 (blue), and 0.3-4.6 million years ago within the Eurasian and North American varieties of J. communis.
These results are biogeographically plausible. The data presented by Mao et al. (2010) indicate that the common Cupressus-Juniperus ancestor was in Asia during the Cretaceous-Tertiary transition. Diversification of Juniperus sections occurred during Paleocene and Eocene time. A period of relatively rapid speciation apparently occurred during the cooling of the Eocene-Oligocene transition, at which time conifers showed substantial poleward range increases (Farjon 2005). Rapid speciation occurred again near the Oligocene-Miocene transition and again during the late Miocene through the Pliocene. These events also coincided with variable climate regimes, and also with the rise of the Qinghai-Tibetan Plateau in Asia and the development of the Basin & Range region in North America and the Central Volcanic Plateau in Mexico. These areas are all current hotspots of juniper diversity. On a global scale, the Pliocene was a period of cooling and drying, producing a rapid increase in the area of semiarid regions, juniper's principal habitat. J. communis, the most widespread of all junipers, favors wetter, cold habitats, which have become extremely widespread in the past 2 million years (the Pleistocene). The data presented by Mao et al. (2010) indicate that the radiation of this species has been relatively recent, within this time frame.
Finally, Mao et al. (2010) note that ancestral Juniperus seem to have inhabited a warm climate with a distribution that coincided with the Madrean-Tethyan Flora of Axelrod (1975), a warm-dry sclerophyllous vegetation type located in the northern semiarid belt of the Eocene, which ran from California to Newfoundland to Spain to Iraq at a time when the Atlantic Ocean was still quite narrow. They suggest that the principal clades of section Sabina may have diverged within this vegetation type, subsequently going extinct in Europe during widespread late Tertiary extinctions in that region, while persisting in Asia and North America. Specifically, Clade 2 could have arisen in North America at this time, with Clade 3 later colonizing North America during Miocene time via the Bering Land Bridge.
Structure of the cladogram, beyond discrimination of the six clades, is generally unclear. This absence of detail is due to several causes:
- Juniperus is a diverse taxon that is actively experiencing speciation in some areas, notably semiarid North America and the Tibetan Plateau. As discussed in the Taxonomic notes for Pinus, processes such as incomplete gene sorting and reticulate evolution complicate understanding the phylogeny of such a genus, and to some degree make a cladogram an inappropriate means of depicting and understanding that phylogeny.
- Most studies of Juniperus have relied upon a proxy form of molecular data, such as essential oil analysis, or have used a single DNA source, such as the ribosomes. Other complex coniferous genera have yielded differing evolutionary information when different plastid sources (ribosomal, nuclear, mitochondrial, or chloroplast) have been studied; see Cupressus and Pinus for further discussion. This level of detailed analysis has not yet been completed for most species of Juniperus.
- Studies to date have relied upon, generally, few and different sources of plant material for their molecular analyses. Comparable studies in taxa using material from many specimens, over a wide area of the species' range, have shown that potentially there is great variability within a single recognized taxon. Some closely-studied Juniperus taxa, most notably J. communis and J. deppeana, have already revealed such complexity. It may be more widespread.
Description
Evergreen shrubs or trees. Branchlets terete, 4-6 angled, variously oriented, but not in flattened sprays (except in J. flaccida). Leaves in decussate (alternating) opposite pairs in 4 ranks or in alternating whorls of 3 in 6 ranks, rarely in whorls of 4 in 8 ranks. Adult leaves closely appressed to divergent, scalelike to subulate, free portion nil to 25 mm (to 32 mm recorded in J. formosana); abaxial gland visible or not, elongate to hemispheric (J. ashei), sometimes exuding white crystalline deposit. Pollen cones with 3-7 pairs or trios of sporophylls, each sporophyll with 2-8 pollen sacs. Seed cones maturing in 1 or 2 years, globose to ovoid and berrylike, 3-20 mm (to 25 mm in J. drupacea), remaining closed, often glaucous; scales persistent, 1-5 pairs or whorls of three, peltate or valvate, tightly coalesced and fused together, thick and fleshy or fibrous to obscurely woody; some sweet (e.g. J. deppeana), many bitter and/or resinous. Seeds 1-3 per scale, round to faceted, wingless; cotyledons 2-6. Seed dispersal by frugivorous birds, which swallow the cones whole, digest the fleshy scales and pass the hard-shelled seeds undamaged through the gut; the bitter taste of many species may be related to discouraging mammalian predators of the seeds. x = 11, usually 2n with 14 taxa at 4n and one at 6n.
Dioecious. Leaves as in sect. Juniperus, including basal abscission zone, except broader (2-3.5 mm wide); 10-25 mm long. Winter buds present. Scale-leaves absent, except for 3-6 whorls of three on the cone peduncles, and 2-3 whorls as bud scales on the winter buds. Dioecious, with male and female cones on different trees. Cones axillary on shoot, on 2-7 mm peduncles; peduncle scale-leaves 1.5-3.5 mm long. Mature female cones large, 18-25 mm, scales valvate in (2-)3(-4) whorls of three, only one whorl fertile, with 1 seed on each fertile scale, the three seeds fused together in a hard bony-textured nut 10-12 mm long; mature in c.18 months from pollination. Type (and only species) J. drupacea Labill. The section is closely allied to sect. Juniperus and may be better treated as a subsection within it.
Dioecious. Leaves not decurrent down stem, with basal abscission zone; in six rows in alternating whorls of three, mostly of 1 kind, subulate, spreading, 5-25(32) mm long, 1-2 mm wide. Winter buds present. Scale-leaves absent, except for 2 (-3) whorls of three on the cone peduncles, and 1-2 (-3) whorls as bud scales on the winter buds. Dioecious, with male and female cones on different trees. Cones axillary on shoot, on a very short 0.3-1 mm peduncle (appearing sessile); peduncle scale-leaves 1 mm. Mature female cones small to medium, 6-15 mm, scales valvate in 1(-2) whorls of three, one whorl fertile, with 1 seed on each fertile scale, the three seeds not fused together; mature in c.18 months from pollination.
About 7-9 species; type J. communis L. The section is divisible into two subsections, a northern group with blue-black mature cones and one broad leaf stomatal band (J. communis and allies; 2-4 spp.), and a southern group with orange-red mature cones and two narrow leaf stomatal bands separated by the midrib (J. oxycedrus and allies; 5 spp.).
Monoecious or dioecious. Leaves decurrent down stem, without basal abscission zone, in four rows in decussate opposite pairs or (occasionally) in six rows in alternating whorls of 3 or eight rows in alternating whorls of 4, of three kinds: scale-like adult leaves on slow-growing and fertile shoots, 1-2 mm, adpressed or with spreading apex, free portion 0-1 mm; whip leaves on strong-growing lead shoots, subulate, spreading, free portion 0.5-2 mm; and juvenile leaves, subulate, spreading, free portion 2-10(-13) mm. In all species, juvenile leaves are produced early in life and often also later on shaded shoots; in a few species (J. pingii, J. procumbens, J. recurva, J. squamata), juvenile-type leaves borne throughout life, with scale-leaves confined to cone peduncles. These and juvenile plants of other species may be distinguished from sect. Juniperus by the leaves being decurrent and lacking a basal abscission zone. Winter buds absent. Monoecious or dioecious, with male and female cones on the same or different trees depending on species. Cones terminal on 1-5(-20) mm scale-leaved side shoots. Mature female cones small to medium, 4-18(-20) mm, scales peltate in 2-4(-5) decussate pairs, 1-4(-6) scales fertile, with 1-3 seeds per fertile scale, the total 1-12+ seeds not fused together; mature in (4-)6-18 months (possibly longer in some spp.) from pollination. About 40 species; type J. sabina L. The section is divisible into several groups based on phenology, cone characteristics and leaf margin form; precise alliances are not yet determined.
Source: Adams (1993, 2018), M.P. Frankis personal observations).
Distribution and Ecology
Primarily Northern Hemisphere, one species (J. procera) in E Africa to 18°S. In many semiarid regions, such as through much of the western USA, northern Mexico and central & southwest Asia, it provides the dominant forest cover on large sections of the landscape. Sect. Juniperus is primarily Eurasian, with one species (J. communis) holarctic, the only member of the section in N. America and by far the most widespread single conifer species. Sect. Caryocedrus is a local endemic in SW Asia and SE Europe. Sect. Sabina occupies most of the range of the genus, except for Eurasia north of 50°N in Europe and 60°N in Asia.
Juniperus is primarily dispersed by birds. As in most Podocarpaceae and Taxaceae, the female cone has been modified to resemble a fruit which appeals to birds and provides significant nutritional value, with the undigested (but somewhat scarified) seed excreted along with a measure of nitrogen that may help to nourish the seed. A few mammals, notably canids (e.g. coyote, fox), and harvester ants have also been shown to consume the fruits and transport the seeds. The phenomenon has been identified in many Juniperus species; Adams and Thornburg (2010) provide a review.
Being bird-dispersed, species of Juniperus are much less affected by water or wind barriers than are most species of the Cupressaceae or Pinaceae. J. bermudiana is endemic to Bermuda and appears to have arisen within the past 1 million years, and about the same time frame applies to J. gracilior of Cuba and Hispaniola, and J. barbadensis of Santa Lucia, Haiti, Jamaica and Cuba. Africa was an island when J. procera differentiated about 42 million years ago, and it is still the only juniper in sub-Saharan Africa. There are many examples of islands where a juniper is the sole conifer. This is also true of sky islands, i.e. very isolated high mountain habitats.
Remarkable Specimens
Available data indicate the largest specimen is found in Juniperus grandis, and the tallest in J. drupacea. There are no age data for most species, but ages exceeding 2000 years have been established for J. grandis and J. przewalskii, and ages exceeding 1000 years for J. scopulorum and J. occidentalis.
Ethnobotany
Numerous cultivars of Juniperus species are widely used for landscaping (Adams 1993). Because the genus is widely distributed in semiarid regions (it grows particularly well on calcareous soils) and some are not particularly palatable to domestic goats, it often affords the only tree of size on the landscape, thus providing an important source of wood for construction, fuel and other domestic uses. The wood is fragrant, usually reddish or reddish-brown, easily worked, very durable, and rarely injured by insects (Dallimore et al. 1967). Its resistance to decay makes it particularly useful for fenceposts and other ground-contact applications. However, it seldom achieves the size or straight grain needed in lumber. Many native peoples have used the aromatic foliage and resins for medicinal or spiritual purposes. "An essential oil is obtained by distillation from wood and leaves. That from the wood is often used for perfumery, sometimes in medicine. Oil from the leaves and shoots is also used in medicine. They have powerful diuretic properties and stock should not be allowed to eat branches" (Dallimore et al. 1967).
Wood and/or foliage are often burned for incense in Buddhist temples.
Cones of J. communis are used for flavoring gin, as are cones of several other species, as well as to flavor more obscure beverages.
A variety of species have proven useful in dendrochronology, delivering dendroclimatic records of up to 1000 years and providing archeological data in the American southwest. However, some species lack circuit uniformity or have a large fraction of missing rings.
Observations
See the respective species. Please inform me if you know of any arboreta with exceptional Juniperus collections.
Remarks
Juniperus is an old Latin name used by Virgil and Pliny (Weber 1987).
"Mutants, or 'sports,' affecting plant habit and foliage are present in all species and are likely related to single-gene mutations. Many have been given formal names or incorrectly ascribed to hybridization. Gymnocarpy (bare seeds protruding from the cone), caused by insect larvae, is occasionally found in most junipers, particularly in the southwestern United States. Specimens with such aberrations may be almost impossible to identify without chemical data" (Adams 1993).
Adams and Mataraci (2011) report: "Adams (1982) found that J. scopulorum var. columnaris Fassett growing near a burning coal seam had columnar shapes. The coal seam has been burning since about 1880. Murphy and Holden (1979) propagated 25 columnar trees from the site and grew them in a smoke-free area. None of the cuttings produced columnar trees, but rather the typical, pyramidal trees of J. scopulorum. They concluded that ethylene from the burning coal induced the columnar shape. But, of course, other gasses from the burning coal (SO2, NO and CO) may affect plant growth. Columnar trees of Juniperus scopulorum are also found downwind of coal burning plants, Butte, MT." They report a similar phenomenon in Juniperus deltoides, also in an area near coal burning.
Citations
Adams, R. P. 1982. The effects of gases from a burning coal seam on morphological and terpenoid characters in Juniperus scopulorum (Cupressaceae). Southwestern Naturalist 27:279-286.
Adams, Robert P. 1993. Juniperus. Flora of North America Editorial Committee (eds.): Flora of North America North of Mexico, Vol. 2. Oxford University Press. This document is available online. Go to http://www.efloras.org, click on "Flora of North America," and search for "Juniperus."
Adams, Robert P. 2014. Junipers of the World: The Genus Juniperus. Fourth edition. Trafford Publishing. Brief versions of the descriptions are available online at Adam's website, www.juniperus.org.
Adams, Robert P. 2018. Evolution of dioecious/monecious taxa in Juniperus, contrasted with Cupressus, Hesperocyparis, Callitropsis and Xanthocyparis (Cupressaceae). Phytologia 100(4):248-255.
Adams, R. P., and T. Mataraci. 2011. Taxonomy of Juniperus deltoides forma yaltirikiana in Turkey: leaf terpenoids and SNPS from nrDNA and petN. Phytologia 93(3):293-303.
Axelrod, D.I. 1975. Evolution and biogeography of Madrean-Tethyan sclerophyll vegetation. Annals of the Missouri Botanical Garden 62:280–334.
Farhat, P., O. Hidalgo, T. Robert, S. Siljak-Yakovlev, I. J. Leitch, R. P. Adams, and M. Bou Dagher-Kharrat. 2019. Polyploidy in the conifer genus Juniperus: an unexpectedly high rate. Frontiers in Plant Science 10, doi: 10.3389/fpls.2019.00676.
Le Genévrier Thurifère. 2009. http://www.multimania.com/thurifere/" target="_blank">Le Genévrier Thurifère, accessed 2009.11.08, now defunct.
Leslie, A. B., J. M. Beaulieu, H. S. Rai, P. R. Crane, M. J. Donoghue, and S. Mathews. 2012. Hemisphere-scale differences in conifer evolutionary dynamics. Proceedings of the National Academy of Sciences 109(40):16217–16221. Note that the cladistic analysis of Juniperus appears only in the Supplement.
Murphy, J. M. and D. J. Holden. 1979. The ethylene from burning lignite as a probable cause of columnarity in North Dakota junipers. Prairie Naturalist 11:11-22.
Yang, Yong, David Kay Ferguson, Bing Liu, Kang-Shan Mao, Lian-Ming Gao, Shou-Zhou Zhang, Tao Wan, Keith Rushforth, and Zhi-Xiang Zhang. 2023. Recent advances on phylogenomics of gymnosperms and a new classification. Plant Diversity 44(4):340–350. https://doi.org/10.1016/j.pld.2022.05.003.
See also
Adams, Robert P. 1969. Chemosystematic and numerical studies in natural populations of Juniperus. Ph.D. dissertation, University of Texas.
Adams, R. P., and A. E. Schwarzbach. 2012. Taxonomy of the turbinate shaped seed cone taxa of Juniperus, section Sabina: sequence analysis of nrDNA and four cpDNA regions. Phytologia 94(3):388-403.
Adams, R. P., and A. E. Schwarzbach. 2013. Taxonomy of the turbinate shaped seed cone taxa of Juniperus, section Sabina: Revisited. Phytologia 95:122–124.
Adams, R. P. and D. Thornburg. 2010. Seed dispersal in Juniperus: A review. Phytologia 92(3):424-434. Available at www.phytologia.org.
Adams, R. P. and T. A. Zanoni. 1979. The distribution, synonymy, and taxonomy of three junipers of the southwest United States and northern Mexico. Southwest Naturalist 24:323-330.
Adams, R. P., E. von Rudloff and L. Hogge. 1983. Chemosystematic studies of the western North American junipers based on their volatile oils. Biochem. Syst. & Ecology 11:85-89.
Farjon (2005) provides a detailed account, with keys and a cladistic tree.
Fassett 1945.
Hall, M. T. 1952. Variation and hybridization in Juniperus. Annals of the Missouri Botanical Garden 39:1-64.
Van Haverbeke 1968.
Vasek 1966.
Zanoni 1978.
Zanoni and Adams 1979.