Araucariaceae
Henkel & W. Hochstetter (1865)
Common names
Araucaria family.
Taxonomic notes
Three genera with 41 species. The genera include:
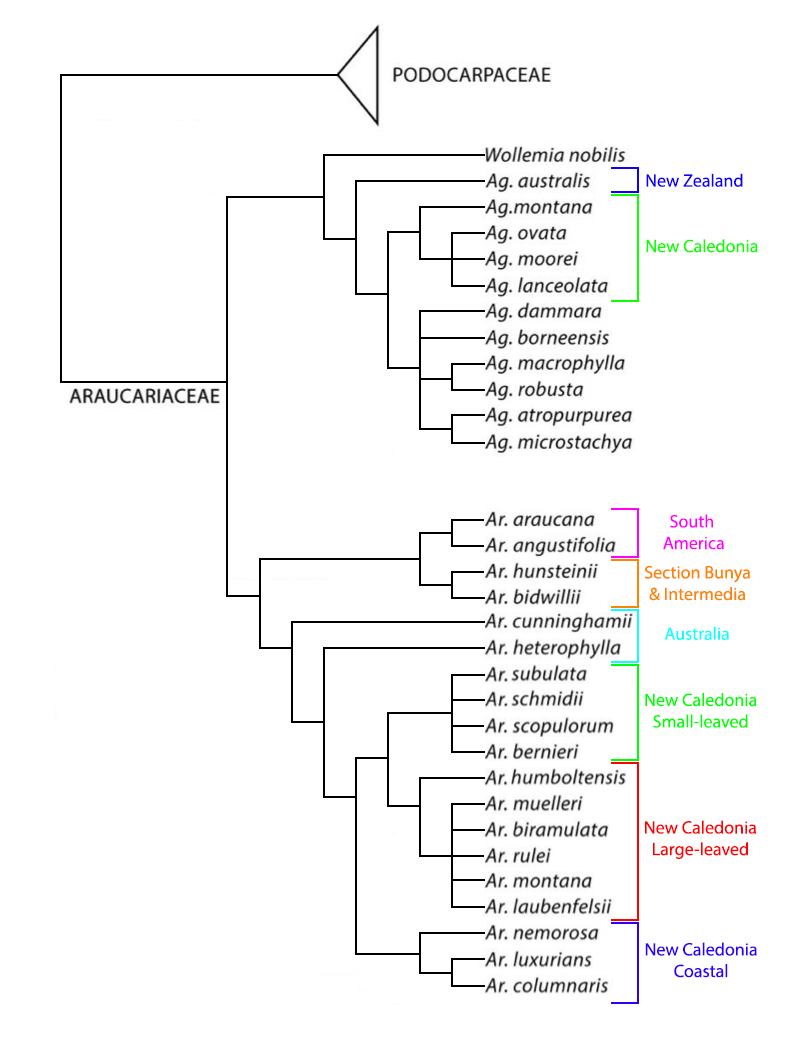
Some authors elevate the family to the level of an order, Araucariales Heintze (1927), which includes the Araucariaceae and the Podocarpaceae. Relationships within the family were long contested, but were finally elucidated by genetic analyses (Setoguchi et al. 1998, Gadeul et al. 2012, Escapa and Catalano 2013). Phylogenetic trees for these analyses agree that all three genera are monophyletic and that Wollemia, sister to Agathis, is the most primitive of the three. Within Araucaria, the four sections previously recognized (Araucaria, Bunya, Eutacta, and Intermedia) are moderately supported, but the close relationship between sections Bunya and Intermedia indicates that Intermedia can be reduced to synonymy, such that section Bunya includes Araucaria bidwillii and A. hunsteinii. Relationships within Agathis seem to have been reasonably well elucidated; A. australis of New Zealand is basal, and the four New Caledonian species are sister to a clade containing all remaining species. "The New Caledonian Araucaria and Agathis species each formed a monophyletic group with very low differentiation in rbcL sequences among them, indicating rapid adaptive radiation to new edaphic conditions, i.e., ultramafic soils, in the post-Eocene era" (Setoguchi et al. 1998). A possible phylogenetic tree is here reproduced, modified from Escapa and Catalano (2013) with added information on the New Caledonia Araucaria from Gadeul et al. (2012); note that one new species, Araucaria goroensis, has since been described in Section Eutacta, where it will fall into the clade of New Caledonia large-leaved species.
Description
Evergreen trees with spirally arranged, narrow or broad leaves often with parallel veins. Dioecious or monoecious. Male cones relatively large, cylindrical, with numerous sporophylls and with ±12 inverted pollen sacs; pollen grains wingless. Female cone usually borne erect, subglobose to ovoid, maturing in two years, relatively large and milky, falling upon maturity; scales one-seeded, without distinct bracts. Cotyledons 4, often fused into 2 double cotyledons (Silba 1986).
Species in the Araucariaceae are endomycorrhizal (McKenzie et al. 2002).
Distribution and Ecology
Primarily the southern hemisphere. Araucarian species are almost all found in rainforest, the principal exceptions being the montane Araucaria araucana of South America and the Araucaria species found in sclerophyllous maquis vegetation on ultramafic substrates in New Caledonia (Kershaw and Wagstaff 2001). In most settings they are among the largest and tallest trees found in their respective forests, despite which they are usually unable to regenerate under a dense canopy in the absence of disturbance, and thus are early seral species.
Worldwide distribution of Araucariaceae, generalized to 1x1 degree cells; cells are color-coded by genus; data from the BRAHMS database at Conifers of the World, accessed 2021.11.11.
"Members of the Araucariaceae are now restricted to the South American and Southwest Asia-Western Pacific region despite their extensive distribution in both hemispheres during the Mesozoic. A relictual status is also suggested by generally low species diversity and disjunct distributions on continental masses. However, relatively high diversity is recorded on islands in the Pacific and the Southeast Asian region, indicating conditions suitable for survival and continuous evolution through a long period of geological time or more recent dispersal and rapid evolution from continental sources" (Kershaw and Wagstaff 2001). This discussion leads on to an intriguing hypothesis: that most, if not all, of the extant Araucarian taxa have evolved since early Tertiary time, and that this evolution and adaptive radiation occurred therefore in the islands of Australasia, where nearly all of the extant diversity (excepting only the two South American species of Araucaria, and two Agathis species on the Malay peninsula) is found. This is a striking pattern because it means that this family of conifers has been most successful in precisely the environment where angiosperms are generally thought to have most successfully replaced conifers - in the tropical rainforests. Several things help to explain this condition, among them the ability of conifers to disperse to and across islands subject to frequent, destructive volcanic disturbance; the trees' longevity, allowing them to wait out the intervals between severe disturbance; and the emergent habit, which diminishes direct competition with the angiosperms. I find these explanations unsatisfactory, though, because they do not explain conifer dominance in locales where many thousands of years pass between highly destructive disturbances, or the persistence of conifers that do not show the emergent habit, or the persistence of conifers in forests where many angiosperms grow as emergents.
In their discussion of the Quaternary history of the Araucariaceae, Kershaw and Wagstaff (2001) present a variety of evidence suggesting that a principal control on Araucarian abundance is fire frequency, with widespread declines in Araucarians during periods of high fire frequency associated with aboriginal burning or, earlier, with episodes of dry climate and high lightning ignition frequencies. Conversely, during periods of low fire frequency and generally wet climate, the Araucarians prosper and become widespread forest dominants. Thus I infer that the Australasian success of Araucarians may be governed more by their ability to adapt to their physical environment - precipitation, fire, edaphic factors - than by their ability to compete effectively with angiosperms. This is a pattern we see repeated commonly in conifers throughout the northern hemisphere: they thrive best in settings where there greatest challenges are not biotic (i.e., competition, herbivory or disease), but abiotic (drought, cold, fire, or poor soils).
A concise, current review of the ecology of the genus is provided by Waters et al. (2002), pages 8-10.
The Araucariaceae are, in general, very much of conservation concern. The IUCN (2020) has identified 4 taxa as critically endangered, 12 as endangered, and 10 as vulnerable, thus 70% of all species in the family are at risk, a greater proportion than in any other conifer family. Principal factors of decline include habitat conversion through agriculture or development, timber harvest, grazing, and climate change.
Remarkable Specimens
The largest trees are in Agathis australis. The only conifer species known to grow larger are Sequoia and Sequoiadendron in the Cupressaceae.
The oldest trees include credible reports of Agathis australis and Araucaria araucana exceeding 1000 years.
Ethnobotany
Some species are important timber trees. Most species of Agathis, for example, are among the largest trees known in the areas where they grow, and those species of Araucaria not native to New Caledonia also reach prodigious size. They have been principal species of concern for logging operations in Australia, New Zealand, and many parts of Malesia. Consequently, many of the finest stands of these species have been logged, and many of the species are threatened or endangered (WCMC 2001). Some species, notably Araucaria bidwillii, are also an important food source.
Limited dendrochronological studies have been done on most of the temperate Australian and New Zealand species. The tropical species have seen little work, and what has been done, indicates that they do not reliably produce annual rings.
Observations
The Araucariaceae include several hardy and attractive species that are very widely used as ornamentals, including Araucaria angustifolia, A. araucana, A. bidwillii and A. heterophylla. These are found in hundreds of botanic gardens and arboreta, and in fact A. heterophylla can be found in even the coldest climates thanks to its popularity as an indoor house plant. Conversely, most of the remaining species of the family are tropical and are not widely known outside of their native ranges. In many cases those ranges are relatively large and the trees fairly easy to locate, but several taxa including Wollemia nobilis, several New Caledonian species of Araucaria, and several Malesian species of Agathis are narrow endemics, endangered by habitat loss.
Remarks
Although the oldest conifers have some araucarioid characteristics, the earliest fossils that can definitely be assigned to the Araucariaceae are of Jurassic, Pliensbachian age (190-183 million years ago. These represent an extinct species of Araucaria (Arrondo and Petriella 1980 in Rothwell et al. 2012). The family soon "expanded and diversified in both hemispheres in the Jurassic and Early Cretaceous and remained a significant component of Gondwanan vegetation until the latter part of the Cenozoic. The development of angiosperms in the Middle Cretaceous probably assisted in the demise of some araucarian components but there was also evolution of new genera. Recorded diversity in the early Cenozoic of Australia is as high as it was in the Early Cretaceous. Continental separation and associated climatic drying, cooling, and increased variability progressively reduced the ranges of conifers to moist, predominantly mesothermal climates on continents. However, tectonic and volcanic activity, partially associated with Australia's collision with Southeast Asia, provided new opportunities for some araucarian components on Asia-Pacific islands." (Kershaw and Wagstaff 2001).
Agathis "is only known from the Southern Hemisphere, beginning in the Cretaceous. Cuticle studies of extant araucarians have enabled accurate comparisons of fossil leaves to living taxa. Araucaria Section Bunya is represented by cones of several types in the Jurassic. In addition to these remains, a suite of araucarian cones showing affinities to several sections of the genus Araucaria have been described from England, Japan and North America" (Stockey 1994).
Citations
Arrondo, O. G., and B. Petriella. 1980. Alicurá, nueva localidad
plantífera Liásica, de la provincia de Neuquén, Argentina.
Ameghiniana 17:200-215.
Escapa, I. H., and S. A. Catalano. 2013. Phylogenetic analysis of Araucariaceae: Integrating molecules, morphology, and fossils. International Journal of Plant Sciences 174(8):1153–1170.
Gaudeul, M., G. Rouhan, M. F. Gardner, and P. M. Hollingsworth. 2012. AFLP markers provide insights into the evolutionary relationships and diversification of New Caledonian Araucaria species (Araucariaceae). American Journal of Botany 99(1):68–81.
Henkel, J. B. and W. C. Hochstetter. 1865. Synopsis der Nadelhöser... xvii, 1. Available: Biodiversity Heritage Library, accessed 2023.03.04.
IUCN. 2020. IUCN Red List version 2020-1: Table 4b: Red List Category summary for all plant classes and families. https://nc.iucnredlist.org/redlist/content/attachment_files/2020_1_RL_Stats_Table_4b.pdf, accessed 2022.09.23.
Kershaw, Peter and Barbara Wagstaff. 2001. The southern conifer family Araucariaceae: history, status, and value for paleoenvironmental reconstruction. Annual Review of Ecology and Systematics 32: 397-414.
McKenzie, E.H.C., P.K. Buchanan, and P.R. Johnston. 2002. Checklist of fungi on kauri (Agathis australis) in New Zealand. New Zealand Journal of Botany 40:269-296.
Rothwell, G. W., G. Mapes, R. A. Stockey, and J. Hilton. 2012. The seed cone Eathiestrobus gen. nov.: fossil evidence for a Jurassic origin of Pinaceae. American Journal of Botany 99(4):708-720.
Royal Botanic Gardens Sydney. 1999. Wollemi. http://www.rbgsyd.nsw.gov.au/html/Wollemi.html, accessed 2001.07.19, now defunct.
Waters, T., C.A. Galley, R. Palmer, S.T. Turvey, and N.M. Wilkinson. 2002. Report of the Oxford University Expedition to New Caledonia. users.ox.ac.uk/~scat1055/expedition/report.htm, accessed 2004.11.20, now defunct.
[WCMC] World Conservation Monitoring Centre. 2001. World Conservation Monitoring Centre - Trees. http://www.wcmc.org.uk/cgi-bin/SaCGI.cgi/trees.exe, accessed 2001.07.13, now defunct.
See also
An International Araucariaceae Symposium was held in Auckland, New Zealand on 14-17 March 2002. Sixty-six papers were presented, subsequently published as Araucariaceae: proceedings of the 2002 Araucariaceae Symposium, Araucaria-Agathis-Wollemia, International Dendrology Society, Auckland, New Zealand, 14-17 March 2002; many are of long-term interest.
Calder, M.G. 1953. A coniferous petrified forest in Patagonia. Bulletin of British Museum (Natural History) Geology 2: 99-137.
Cantrill, D.J. 1992. Araucarian foliage from the Lower Cretaceous of southern Victoria, Australia. International Journal of Plant Science 153: 622-645.
Cookson, I.C. and S.L. Duigan. 1951. Tertiary Araucariaceae from southeastern Australia, with notes on living species. Australian Journal of Scientific Research Ser. B 4: 415-449.
Enright and Hill 1990.
Enright, N.J.; Ogden, J.; Rigg, L.S. 1999. Dynamics of forests with Araucariaceae in the western Pacific. Journal of Vegetation Science 10(6):793-804.
Florin, R. 1963. The distribution of conifer and taxad genera in time and space. Acta Horti Bergiana 20: 122-312.
Hill, R. S. and A. J. Bigwood. 1987. Tertiary gymnosperms from Tasmania: Araucariaceae. Alcheringa 11: 325-335.
Hill, R.S. and T.J. Brodribb. 1999. Southern conifers in time and space. Australian Journal of Botany 47: 639-696.
de Laubenfels (1972).
Miller, C.N. 1977. Mesozoic conifers. Botanical Review 43: 217-280.
Miller, C.N. 1988. The origin of modern conifer families. P. 448-486 in C.B. Beck (ed.), Origin and evolution of gymnosperms. Columbia
University Press, New York, NY.
Pole, M.S. 1994. The New Zealand flora - entirely long-distance dispersal? Journal of Biogeography 21: 625-635.
Macphail, M. K. 1997. Comment on M. Pole (1994): "Is New Zealand flora - entirely long distance dispersal?" Journal of Biogeography 24: 113-117.
Raven, P.H. and D.I. Axelrod. 1972. Plate tectonics and Australasian palaeobiography. Science 176: 1379-1386.
Raven, P.H. and D.I. Axelrod. 1974. Angiosperm biogeography and past continental movements. Annals of the Missouri Botanical Garden 61: 539-673.
Stockey, R.A. 1982. The Araucariaceae: an evolutionary perspective. Review of Palaeobotany and Palynology 37: 133-154.
Stockey, R.A. 1990. Antarctic and Gondwana conifers. P. 179-191 in Antarctic Paleobiology (T.N. Taylor and E.L. Taylor, eds.). New York: Springer-Verlag.
Stockey, R.A. 1980b. Jurassic araucarian cone from southern England. Palaeontology 23: 657-666.
Stockey, R.A. 1994. Mesozoic Araucariaceae: morphology and systematic relationships. Journal of Plant Research 107: 493-502.
Stockey, R.A., M. Nishida, and H. Nishida. 1990. Structure and diversity of the woody conifer seedling-like structures from the upper Cretaceous of Hokkaido, Japan. Botanical Gazette 151: 252-262.