The peculiar foliage morphology that gave Retrophyllum its name (Jean.claude at Wikimedia Commons, 2012.02.18).

Retrophyllum minus in its native aquatic habitat, New Caledonoia [Adam Black, 2020.02.01, Facebook post].

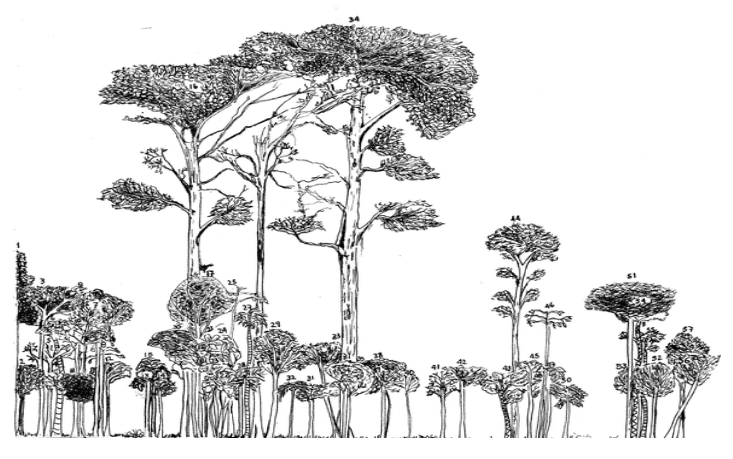
Forest sectional illustration showing emergent podocarps (chiefly Retrophyllum rospigliosii) in primary Andean rain forest.

Illustration of Retrophyllum vitiense from Seemann's Flora Vitiensis, available at the Biodiversity Heritage Library, accessed 2021.11.17.

Retrophyllum
Page 1989
Common names
Taxonomic notes
A genus of six species:
All species except the recently-recognized R. filicifolium were part of Nageia until Page (1989) segregated them. At the time, the segregation seemed arbitrary and was unsupported by any statement of rationale. Herbert et al. (2002) used a combination of anatomical characters and molecular methods including trnL-F sequences, chloroplast RFLPs and RAPDs to compare the various species of Retrophyllum to their sister taxa in the Podocarpaceae. Their results indicate the genus is apparently monophyletic, and is sister to Nageia and Afrocarpus. Within the genus, there is a clear correspondence between geography and phylogeny: the two New Caledonian species, R. comptonii and R. minus, are most closely related; next is the Fijian species R. vitiense; and these three form a clade sister to one of the South American species, R. rospigliosii. Another species, R. filicifolium, was segregated from R. vitiense after Herbert et al. did their work (Mill 2016). The second South American species, R. piresii, is extremely rare and Herbert et al. (2002) were unable to extract a usable molecular sample from the one herbarium specimen that they could sample. The one specimen of R. vitiense they sampled was from Fiji, thus, no R. filicifolium were sampled.
The genus received monographic treatment by Mill (2016), who came to similar conclusions, dividing the genus into three groups represented by two Pacific species exclusive of New Caledonia, two New Caledonian species, and two South American species. Mill (2016) also recognized R. filicifolium (which had already been described as Podocarpus filicifolius by Netta Gray in 1962) and precisely described the geographic and morphological basis for separating that species from R. vitiense.
Description
Dioecious trees or shrubs. Bark exfoliating in short vertical strips or small pieces, or as large plates. Vegetative shoots dimorphic with either both foliage and scale leaves or only scale leaves (Pacific and New Caledonian species) or monomorphic with foliage leaves only (South American species), heterofacially flattened at least in juvenile phase, in adult phase sometimes not fully so (New Caledonian species). Terminal buds developed on shoots. Leaves of juvenile phase distichous, heterofacially turned at base so that all leaves along one side of the axis present their abaxial surface to the light and all those along the other side present the adaxial surface; those of the adult phase are either distichous and flattened in 2 ranks (Pacific and South American species) or wholly or partly in 4 ranks that show only minimal heterofacial turning (New Caledonian species). Adult leaves subsessile with bases decurrent and crossing over to a departure point on the opposite side and so creating a zigzag pattern along the branch; blades bifacially flattened, coriaceous, relatively thin (Pacific and South American species) or thick to very thick (New Caledonian species), with or without a visible midrib; resin canals below the vascular bundle; the South American species also have 2 to 6 lateral bundles around the leaf margin. Pollen cones usually lateral but sometimes terminal in R. vitiense, 1–8 together; cylindrical, ellipsoid or ovoid; microsporophylls imbricate. Pollen 2-saccate. Female cones borne on current growth; cones pedunculate; carpophore absent; cone axis with 1 to 8 sterile bracts below the fertile bract. Receptacle vestigial. Ovule inverted. Epimatium present. 2n = 20 (Mill 2016).
Mill (2016) also provides a key to the six species, and as his monograph is an open-access document readily available online, I will not repeat it here.
Distribution and Ecology
Malesia, New Caledonia, Brazil, Venezuela and Colombia (Farjon 1998). Like Lepidothamnus, Pectinopitys, Podocarpus, and Prumnopitys, the genus has representatives on both east and west sides of the Pacific Ocean. It thus predates the breakup of Gondwana, which happened in late Cretaceous time when South America separated from Antarctica.
Remarkable Specimens
R. rospigliosii and R. vitiense attain comparable sizes, and both are considerably larger than the other species. However, data are few.
Ethnobotany
Retrophyllum respigliosii and R. vitiense are exploited for their timber. R. rospigliosii has been explored for use in dendrochronology (Becerra-Montalvo and Zevallos-Pollito 2013).
Observations
See the species accounts.
Remarks
Retrophyllum is from the Latin "retro" (backwards) and "phyllum" (leaf), because the leaves are twisted so that along one side of the stem axis all the abaxial surfaces face uppermost, while along the other side of the axis all the adaxial surfaces are turned uppermost (Mill 2016).
Plants of this genus have been found in early Miocene (about 20 million years ago) sediments in southern New Zealand (Pole 2007).
Citations
Becerra-Montalvo, Vitoly and Percy A. Zevallos-Pollito. 2013. Determinación del turno de corta de Cedrela odorata L., Retrophyllum rospigliosii Pilger, y Prumnopitys harmsiana Pilger a través del estudio dendrocronológico en San Ignacio, Región Cajamarca-Perú. El Ceprosimad 2(1):96-102.
Herbert, J., P.M. Hollingsworth, M.F. Gardner, R.R. Mill, P.I. Thomas, and T. Jaffré. 2002. Conservation genetics and phylogenetics of New Caledonian Retrophyllum (Podocarpaceae) species. New Zealand Journal of Botany 40:175-188.
See also