Abies
Miller 1754
Common names
Fir [English]; نراد [Arabic]; 冷杉属 [Chinese]; Tanne [German]; sapin [French]; abete [Italian]; モミ属 [Japanese]; Пихта [Russian]; abies [Spanish].
Taxonomic notes
Approximately 51 species, making it the fourth-largest conifer genus (after Pinus, Podocarpus and Juniperus). Linnaeus initially (1753) assigned all firs, spruces and pines to the genus Pinus. Miller promptly (1754) assigned firs to the genus Abies (type species: A. alba). Various authors have assigned it to ranks above the level of family: Order Abietales Koehne 1893; Family Abietaceae Bercht. et J. Presl 1820; Subfamily Abietoideae Rich. ex Sweet 1826; Tribe Abieteae Rich. ex Dumort. 1827; and Subtribe Abietinae Eichler 1887. Infrageneric systematics have been a long-standing puzzle, with the current extant classification having maintained much of its structure through revisions by Liu (1971), Rushforth (1987), Farjon (2010), and Xiang et al. (2018). The last of these analyses has made extensive use of molecular evidence, with the chloroplast genome most useful in elucidating the differences between taxa; nonetheless, significant uncertainties remain in cladistic relationships between the sections (primarily, the placement of sect. Bracteata), and there are numerous uncertainties regarding relationships within the sections. The classification shown here is that of Xiang et al. (2018), who also provide formal descriptions of each of the sections.
Several studies have examined molecular evidence from a substantial majority of species in the genus. Semerikova and Semerikov (2014, 2016, 2018) use nuclear (nrITS), chloroplast (cpDNA), and mitochondrial (mtDNA) lines of evidence for 37 species and some infraspecifics. Xiang et al. (2015, 2018) use nrITS, cpDNA, and mtDNA lines of evidence for 52 taxa, and Xiang et al. (2024) use single-copy nuclear genes; this last study addresses various methodological issues and data limitations in the earlier studies, providing the clearest picture to date, but still concludes that relationships within section Pseudopicea, in particular, remain unclear, and that future studies should "incorporate multiple individuals per taxon and more comprehensive genetic information." The studies agree in several important points:
- All analyses support a western North American origin for the genus, very possibly in California; Xiang et al. (2015, 2024) estimate a Paleocene origin with initial dispersal from North America into Asia around the Eocene/Oligocene boundary. Diversification between the East Asian and Mediterranean taxa (Section Pseudopicea and Section Abies) occurred at about the Oligocene/Miocene boundary (Xiang et al. 2024).
- Section Balsamea has an unclear origin. These are species of the Holarctic and thus have been much more biogeographically mobile than species confined to more southerly mountain belts. The nrITS and cpDNA cladograms differ substantially, as noted by Xiang et al. (2015), which they attribute to an incomplete lineage sorting event that selectively lost different mtDNA and cpDNA types in the North American vs. East Asian species of this Section. Xiang et al. (2024) place these species sister to the Abies-Pseudopicea clade, which might be biogeographically plausible if those clades formerly included now-extinct North American taxa; or it might simply be incomplete lineage sorting.
- Sections Amabilis and Nobilis are confounded in the cpDNA analysis of Xiang et al. (2015, 2018), and are nearly sister in that of Semerikova and Semerikov (2016), which did not sample A. procera; in the nrITS analysis Xiang et al. (2015) placed Section Nobilis sister to Section Amabilis, sister to Section Grandis. That is reversed in the analysis of Xiang et al. (2024), which puts Nobilis sister to Bracteata, and Amabilis sister to Grandis. One puzzle is that here, as in almost every other classification, A. amabilis falls into a section with A. mariesii of Japan. Xiang et al. (2015) estimate the date for this disjunction at 11.3 mya and suggest dispersal of the A. mariesii progenitor via Beringia at that time, with its subsequent isolation to Japan.
- The mtDNA results display a biogeographic pattern, different from that shown by the cpDNA and nrITS data, as noted in two studies (Xiang et al. 2015, Semerikova and Semerikov 2016). This occurs because mitochondrial DNA comes from the maternal parent and as such is often a marker for hybridization or introgression. Xiang et al. (2015) note that "The Asian members of sect. Balsamea may have undergone frequent introgression mediated by pollen flow, which resulted in the clustering with sect. Pseudopicea." Similarly, Semerikova and Semerikov (2016) note "This is most clearly represented by the separation of the Asian fir species in the AFLP tree [mtDNA] into northern and southern groups."
The greatest center of diversity for Abies is now in the eastern Himalaya, with secondary concentrations in Japan, the Klamath/Siskiyou region of North America, and around the Mediterranean. The greatest rate of speciation for the East Asian taxa began with the rising of the Himalaya, leading to the high species diversity now found in that area. Abies in general has an association with seasonal rainfall, high moisture, and cool temperatures; thus the rising of the Himalaya in a formerly arid and semiarid region produced a variety of new environments, and the available Abies taxa radiated into those new niches. Conversely, the episodes of mountain-building in western North America had the opposite effect, introducing aridity to formerly mesic landscapes and driving vicariant speciation in the California and Mexico regions by essentially confining Abies to mountaintop "islands" of cool, seasonally wet climate. A similar process was at work in the Mediterranean, where taxonomic diversity is tied to narrow geographic endemism as in the endemic firs of Sicily, Spain, Morocco and Algeria (Xiang et al. 2024).
Several species have closely allied sibling species, e.g., A. balsamea-A. fraseri, A. concolor-A. grandis, and A. magnifica-A. procera. When distributions of species overlap, introgression between the taxa is common if the species are in the same section.
Description
Tree: Like most other genera of Pinaceae, species of Abies are typically monoecious evergreen trees with a spire-like or conic crown that often becomes flattened or rounded in old trees. Near the alpine treeline, trees may grow with reduced stature and a contorted growth form, often forming krummholz at the upper limits of tree growth. Their form differs from most genera of Pinaceae in its exceptional uniformity; Abies typically possess a single straight trunk with regularly spaced branch whorls produced at the rate of one whorl per year, so that it is sometimes possible to determine the age of a tree 50 years old simply by counting branch whorls. The branch pattern is also exceptionally regular, with a single terminal and two lateral shoots produced each year at the tip of most active branches. Thus, Abies displays a geometric regularity of form that is only a little less common in Picea and Larix, is rather unusual in Pinus, and is not found in Tsuga.
Bark: Smooth and thin on young trees, bearing resin blisters, in age often thick and furrowed or flaking in plates. In most species the bark provides little protection against fire, although a few species are fire-tolerant.
Branches: Whorled, spreading, flattened into a horizontal plane, with irregular internodal branches occasionally produced by epicormic sprouting.
Shoots: Short (spur) shoots are absent. Twigs may be grooved or smooth; prominent leaf scars, circular to broadly elliptic, flush with twig surface, are slightly depressed or slightly raised evenly all around.
Leaves: Borne singly, persisting 5 or more years (maximum 53 years in A. amabilis), spirally arranged but often proximally twisted so as to appear either 1-ranked (pointing up like toothbrush bristles) or 2-ranked, sessile, typically constricted and often twisted above the somewhat broadened base, sheath absent; linear-lanceolate, flat, with two white stomatal bands beneath, keeled below, rounded or notched at the apex, with two resin ducts. Since many species are early seral and form dense stands, there is often a conspicuous structural difference between shade foliage, just described, and sun foliage, growing near the top of the crown in full sunlight. Sun foliage is more or less erect, incurved to nearly falcate, thickened or quadrangular. When identifying species, reference is made to mature shade foliage, unless otherwise noted. Buds are ovate or oblong with a rounded or pointed apex; terminal buds are surrounded by 4-5 secondary buds. Cotyledons 4-10.
Pollen cones: Axillary, densely clustered along undersides of current year's twigs, globular or conic, appearing in the spring, pendant, yellow to red, green, blue, or purple, leaving gall-like protuberances after falling.
Seed cones: Borne on year-old twigs, maturing in 1 season, erect, ovoid to cylindric, generally resinous, dehiscent (falling apart on maturity), cone axis persisting as an erect rachis on the branch.
Cone scales: Lacking apophysis and umbo, rounded with a hidden or sometime protruding lobed bract (esp. see A. bracteata).
Seeds: Winged, with a resin sac at the wing-seed juncture.
Wood: Lacks resin ducts.
Sources: Li (1975), Silba (1986), Rushforth (1987).
x=12 (Hunt 1993).
Hunt (1993) advises: "Notes on the following features, made at the time of collection of specimens, are useful in identification.
- "Size and placement of resin canals in the leaves as seen in cross section with a hand lens when a leaf is pulled apart or cut with a sharp knife.
- "Stance of the leaves, e.g., whether they are in flat sprays ('2-ranked') or point up like brush bristles ('1-ranked'), and whether some on a twig point in a direction different from others on the same twig.
- "Differences in color and glaucousness of the lower and upper leaf surfaces.
- "Shape of leaf apex as observed with a hand lens.
- "Distribution of stomates - and number of rows of stomates - on the abaxial and adaxial leaf surfaces, particularly midway between base and apex of leaf.
- "Leaf-scar periderm color. Pull a leaf from a twig and note, with a hand lens, the color of the scar's periphery.
- "Presence or absence of resin on the buds (collect a few extra buds for dissection). If buds are not available (as in the early part of the growing season), collect older branch material bearing old bud scales.
- "Cone color of both pollen and seed cones (binoculars are handy to note this feature of the seed cones)."
Distribution and Ecology
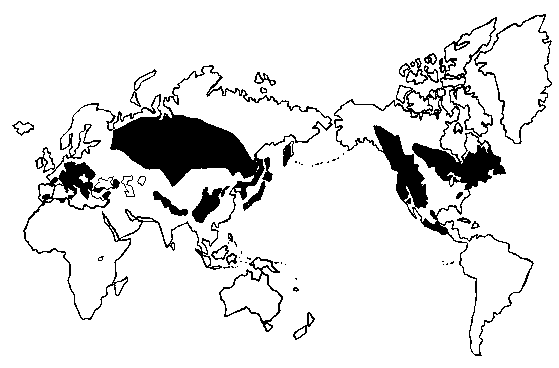
N America, C America, Europe, N Africa, Asia (S to Himalaya, S China, and Taiwan), i.e., temperate and boreal regions of the northern hemisphere, chiefly in mountainous regions (Li 1975). Excepting the two boreal species, A. balsamea (in North America) and A. sibirica (in Eurasia), the genus is confined to mountainous areas in the subtropical and temperate latitudes of the northern hemisphere. Within this realm, its distribution is confined largely according to its ecological requirements and its paleobotanical history.
For information on ecological requirements, paleobotany and modern distribution, see Farjon (1990).
Remarkable Specimens
Few species of Abies exceed 200 cm dbh, 60 m tall, or 500 years old.
The greatest diameter (295 cm dbh) is found in A. magnifica in California, United States. The second-greatest diameter (253 cm dbh) is found in A. procera in Washington, United States. The greatest diameter that can be documented in the eastern hemisphere is found in A. alba; a specimen in Montenegro measured 227 cm dbh. There are sketchy accounts of A. religiosa in Mexico, and A. nephrolepis and A. sachalinensis in east Asia, that might exceed this size. Historical records speak of A. spectabilis in the Himalaya up to 340 cm dbh, but no modern records even approach that size.
The tallest height (83.9 m) is found in A. procera in Washington, United States. The second-tallest Abies is A. chensiensis in Tibet, 83.2 m tall. Given the limited sampling that has been done, it is very possible that taller specimens of A. chensiensis could be found, perhaps exceeding the A. procera record, although, much taller A. procera were recorded historically.
The greatest age, >900 years, is found in A. amabilis in British Columbia, Canada. The second-greatest age, 665 years, is found in A. magnifica var. shastensis in northern California. The oldest in the eastern hemisphere is 660 years for A. forrestii in Sichuan, China.
Ethnobotany
"North American firs are cut for pulpwood and lumber and, largely from plantations, for Christmas trees. They are also grown as ornamentals. Species of Abies frequently have a pleasant odor; their foliage has been used as a stuffing material for pillows. Most commercial products with "pine odors" are in fact scented with essential oils distilled from Abies foliage by Russian farmers" (Hunt 1993).
Many species of Abies have been used in dendrochronology, but they tend to occur on sites with ample moisture and deep soils, where they grow quickly and do not attain great ages. Such conditions characterize closed forest situations, where competition between individuals tends to obscure the effect of large-scale factors, such as climate, on tree growth. Moreover, the tree rings are often difficult to discern and the wood decays quickly after tree death. Consequently, Abies has received relatively little attention in comparison with, say, Pinus or Picea. What work has been done, has largely focused on ecological problems such as stand age structure and forest regeneration after disturbance.
Observations
See the species accounts.
Remarks
"Abies" was the Latin name for an Old World species, probably A. alba but the Romans likely encountered a variety of Mediterranean firs.
Citations
Farjon, Aljos. 1990. Pinaceae: drawings and descriptions of the genera Abies, Cedrus, Pseudolarix, Keteleeria, Nothotsuga, Tsuga, Cathaya, Pseudotsuga, Larix and Picea. Königstein: Koeltz Scientific Books.
Farjon, Aljos. 2010. A Handbook of the World's Conifers. Leiden, Netherlands: Brill Academic Publishers.
Hunt, Richard S. 1993. Abies. Flora of North America Editorial Committee (eds.): Flora of North America North of Mexico, Vol. 2. Oxford University Press.
Li Hui-Lin. 1975. Flora of Taiwan, V.1, parts 1-8. Taipei: Epoch Publishing.
Liu Tang-Shui. 1971. A Monograph of the Genus Abies. Taipei: National Taiwan University.
Miller, P. 1754. The Gardeners Dictionary, abridged 4th edition. V.1, p.11.
Rushforth, K. D. 1987. Conifers. New York: Facts on File. 232p.
Semerikova, S. A., and V. L. Semerikov. 2014. Molecular phylogenetic analysis of the genus Abies (Pinaceae) based on the nucleotide sequence of chloroplast DNA. Russian Journal of Genetics 50:7–19.
Semerikova, S. A., and V. L. Semerikov. 2016. Phylogeny of firs (genus Abies, Pinaceae) based on multilocus nuclear markers (AFLP). Russian Journal of Genetics 52:1164–1175.
Semerikova, S. A., Y. Y. Khrunyk, M. Lascoux, and V. L. Semerikov. 2018. From America to Eurasia: a multigenomes history of the genus Abies. Molecular Phylogenetics and Evolution 125:14–28.
Silba, J. 1986. An international census of the Coniferae. Phytologia memoir no. 8. Corvallis, OR: H.N. Moldenke and A.L. Moldenke.
Xiang, Qiaoping, Ran Wei, Y.-Z. Shao, et al. 2015. Phylogenetic relationships, possible ancient hybridization, and biogeographic history of Abies (Pinaceae) based on data from nuclear, plastid, and mitochondrial genomes. Molecular Phylogenetics and Evolution 82:1–14.
Xiang, Qiaoping, Ran Wei, Yan‐Mei Zhu, et al. 2018. New infrageneric classification of Abies in light of molecular phylogeny and high diversity in western North America. Journal of Systematics and Evolution 56(5):562-72. https://doi.org/10.1111/jse.12458.
Xiang, Qiaoping, Jie Yang, David S. Gernandt, et al. 2024. Ecological and evolutionary factors contribute to the uneven diversification of firs in the northern hemisphere. Journal of Biogeography jbi.15055. https://doi.org/10.1111/jbi.15055.
See also
Farjon, A. and K. D. Rushforth. 1989. A classification of Abies Miller (Pinaceae). Notes of the Royal Botanic Garden Edinburgh 46(1):59-79.