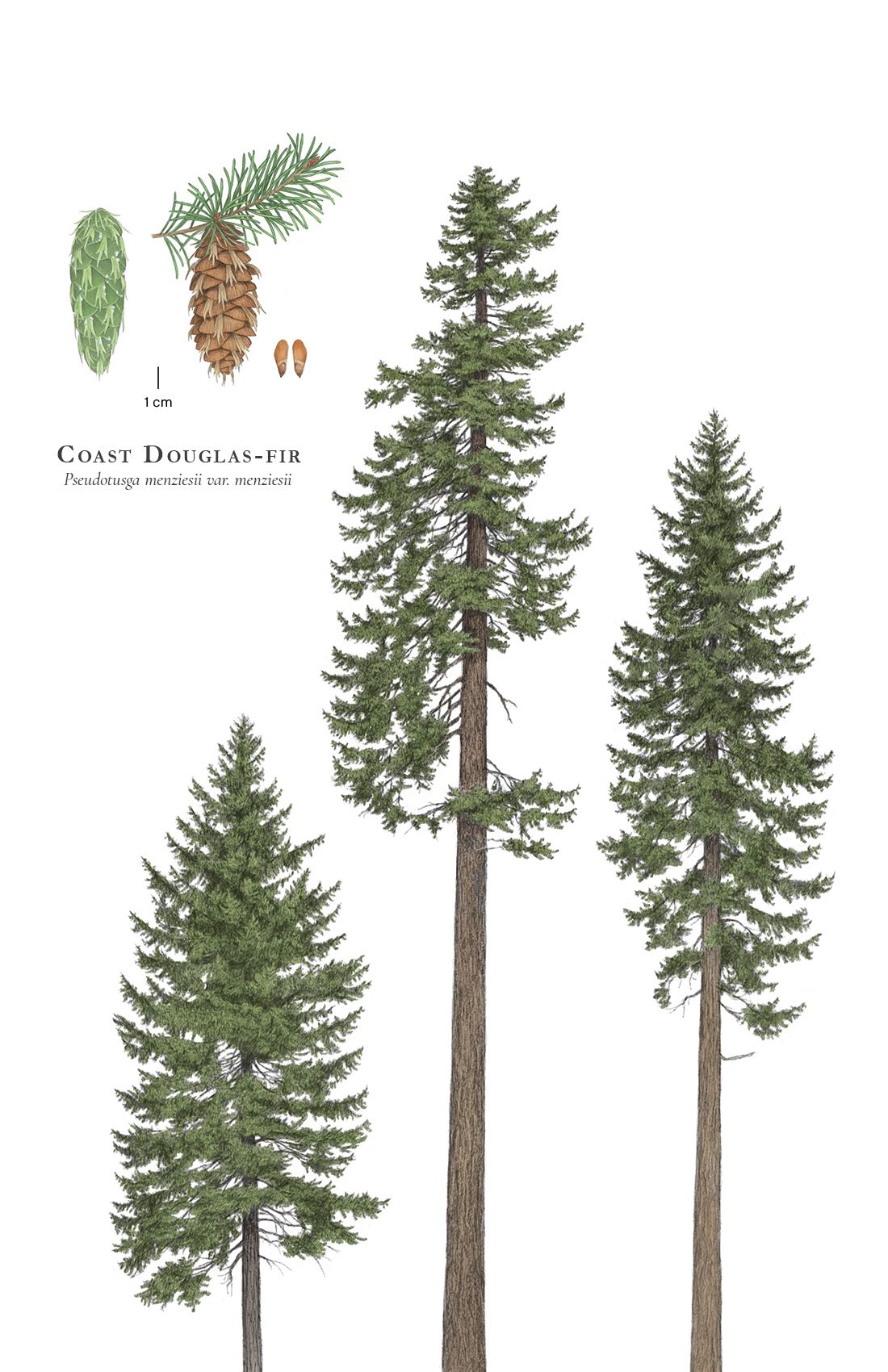
Pseudotsuga menziesii
(Mirbel) Franco 1950
Common names
Douglas-fir; Douglas, yellow or red spruce; Oregon pine, Douglastree (Peattie 1950), sapin de Douglas [French] (Lipscomb 1993); abeto de Douglas, piño Oregon [Spanish] (Burns & Honkala 1990), Douglasia, abete de Douglas, abete americano, pino dell'Oregon [Italian], Douglasie [German], Douglasgran [Danish].
Taxonomic notes
Syn: Abies menziesii Mirbel 1825; Pinus taxifolia Lambert 1803, not Salisbury 1796; Pseudotsuga douglasii (Lindley) Carrière; Pseudotsuga taxifolia (Lambert) Britton. These are only the most prominent of many, many names this species has borne; I don't know of any other species that has been so widely recognized as unique and readily distinguishable, yet has held so many names. An extremely complex nomenclatural controversy surrounded this species from 1800 to 1950; for a detailed recounting of this controversy, see Reveal [n.d.]; for an even more detailed account see Hermann (1982).
There are two intergrading subspecies:
The subspecies are sympatric in southern British Columbia and northeastern Washington (Lipscomb 1993). There is limited evidence for other varieties or subspecies in Mexico. Flous (1934a, 1934b) named an excessively large number of taxa throughout the American range of the genus Pseudotsuga based on cone characters that proved poorly formulated, unreliable and subject to individual variation. Little (1952) subsequently provided a categorical rejection of Flous' interpretation. Snajberk and Zavarin (1976) performed a rangewide analysis of needle terpene compositions that found consistent differences between coastal, Sierra Nevada, northern interior, and southern interior regions. Peng and Adams (1989) presented evidence from Mexico suggesting that (at least some) Douglas-fir in northeast Mexico is genetically distinct from the taxon in North America. The most recent work, by Adams and colleagues (Adams et al. 2012, Adams and Stoehr 2013, Adams et al. 2013) has again examined terpenes, along with leaf essential oils and genetic markers. This work confirmed the status of Ps. macrocarpa as a clearly distinct taxon, and provided new detail on differentiation within Ps. menziesii. These analysis strongly support the traditional division into coastal and interior subspecies, with no significant discontinuities over the long latitudinal range of the species; while the genetic data offer some weak support for the existence of a distinct Oaxacan variety. In general, though, Mexico is more accurately described as the area of greatest genetic diversity within subspecies glauca, rather than as the home of sundry infraspecific taxa.
Description
Trees with a single round trunk to 100 m tall and 440 cm dbh, with a narrow to broadly conic crown that typically becomes flattened or irregular in age; most trees more than about 350 years old have primarily epicormic branches (Van Pelt and Sillett 2008). Bark variable; gray to black or red-brown, generally with longitudinal fissures, scaly, sometimes flaking. Twigs slender, yellowish-green and minutely pubscent, turning gray and glabrous, bearing small circular leaf scars. Leaf buds ovoid, acute, 10 × 5 mm, not resinous. Leaves radially inserted but sometimes appearing 2-ranked on shade foliage; 15-30(-40) × 1.2-1.5 mm, various shades of green (yellow-green to dark-, gray-, or blue-green, usually uniform color on a branch, but adjacent branches on the same tree can have markedly different color), apex obtuse to acute. Pollen cones 15-20 mm long, yellow-red. Seed cones hanging on 5-10 mm long peduncles, ovoid, variably elongate, obtuse or pointed, 4-10 × 3-3.5 cm when opened, green with yellow-green 3-pointed bracts, ripening to gray or golden brown with lighter, straight to reflexed bracts. Seeds 6-8 × 4-5 mm, light brown, with a 9-14 mm long, yellow-brown wing (Lipscomb 1993, Farjon 2010). Detailed information on pollen, including photomicrography, is provided by Davis (1999).
Lipscomb (1993) distinguishes the subspecies as follows:
- Subsp. menziesii has bracts straight, appressed; seed cones 6-10 cm; leaves yellowish green; in the Pacific Coast region.
- Subsp. glauca has bracts spreading, often reflexed; seed cones 4-7 cm; leaves bluish green to dark green or gray-green; in the Rocky Mountain region.
Distribution and Ecology
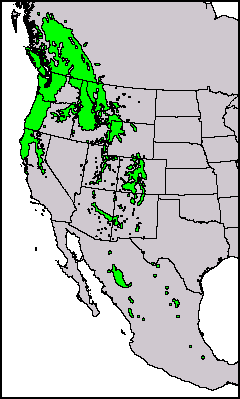
Canada: British Columbia and Alberta; United States: Washington, Oregon, California, Idaho, Montana, Wyoming, Nevada, Utah, Colorado, Arizona, New Mexico, and Texas; Mexico: Chihuahua, Coahuila, Durango, Hidalgo, Nuevo León, Oaxaca, Sonora, and Zacatecas. The coastal subspecies menziesii occurs from central British Columbia south along the Pacific Coast for about 2,200 km to latitude 34° 44'. The interior subspecies glauca is found in the intermountain west, and along the Rocky Mountains and both Sierra Madre chains from Alberta to Oaxaca. Both subspecies tend to occur at progressively higher elevations from north to south. The principal limiting factors are temperature in the north of the range and moisture in the south. Consequently, Douglas-fir is found mainly on southerly slopes in the northern part of its range, and on northerly exposures in the southern part. At high elevations in the southern Rocky Mountains, however, Douglas-fir grows on the sunny slopes and dry rock exposures (Burns and Honkala 1990). See also Thompson et al. (1999).
Native distribution of Pseudotsuga; P. menziesii in red. Distribution data from GBIF, 2021.02.22.
Pseudotsuga menziesii has been planted both as an agroforestry and as an ornamental tree on six continents. It has naturalized in much of Europe (Schmid et al. 2014). It has been reported as invasive in New Zealand, Argentina, and Chile, and in Europe it has the potential to become invasive in Germany, Austria, Bulgaria and Great Britain (Da Ronch et al. 2016).
Remarkable Specimens
The largest trees are found in subsp. menziesii. This is the largest member of the Pinaceae (the second largest, Picea sitchensis, grows with P. menziesii in the coastal forests of NW North America) and larger by far than any other species in the genus.
The tallest tree in each of the following European countries is a Douglas-fir:
- United Kingdom: in the Highlands of Scotland, 68.4 m tall and 139 cm dbh (e-mail Kouta Rasanen 2017.08.16)
- Slovenia: in Savinjska, 67.0 m tall and 470 cm girth, measured in 2020 (Monumental Trees 2022).
- France: in Renaison, 66.6 m tall and 120 cm dbh when measured in 2015 (e-mail Kouta Rasanen 2017.08.16)
- Germany: in Günterstäler Stadtwald, 66.58 m tall and 101 cm dbh (e-mail Kouta Rasanen 2017.08.16)
- Czech Republic: in Železný Brod, 64.05 m tall and 88 cm dbh) (Monumental Trees 2017a)
- Italy: 62.47 m tall and 106 cm dbh (Venturini 2016)
- Ireland: at at Powerscourt House and Gardens, 61.50 m tall and 169 cm dbh when measured in 2013 (Monumental Trees 2013a)
- Ukraine: 63 m tall in 2023 (Facebook post 2023.04.15 by Реєстр Рекордних Дерев in Douglas Fir Trees group).
- Poland: a tree in Lower Silesia, 59.4 m tall and 349 cm girth in 2021 (Monumental Trees 2022).
- Denmark: north of Slåen Sø in East Jutland, 53.7 m tall (direct tape measurement) when measured in 2018 (Christiansen 2018)
- Belgium: 51.20 m tall and 120 cm dbh when measured in 2013 (Monumental Trees 2013b)
- Luxembourg: 51 m tall when measured in 2006 (Monumental Trees 2006)
- Netherlands: 50.40 m tall and 123 cm dbh when measured in 2017 (Monumental Trees 2017b)
All of these trees, of course, were planted, and they substantially exceed native species in height.
Very old trees are recorded in both subspecies. For subsp. menziesii, there's a maximum age of 1350 years, but this was a ring count from the stump of a dead tree. I know of no live-tree ages over 1000 years. However, for subsp. glauca, there is a crossdated age of 1275 years for a living tree in northern New Mexico (more precisely, it was living when it was sampled in the late 1980s or early 1990s, but there is no recent information on its status).
Ethnobotany
Pseudotsuga menziesii is one of the world's most important timber trees (Burns and Honkala 1990, Lipscomb 1993). Data are hard to find, but economically, either this or Pinus radiata is likely the most important softwood on Earth; P. menziesii wood is more valuable, but P. radiata grows faster. It is clearly the most economically important softwood in the U.S. (Oswalt et al. 2017) and in Europe (Pulkrab et al. 2014), where it is the most abundant non-native tree in cultivation (Da Ronch et al. 2016).
Douglas-fir has been a popular selection for the U.S. Capitol Christmas tree, being used 6 times (as of 2020) since the tradition began in 1964. It also makes up the bargain class of U.S. commercial Christmas trees, being cheap and easy to prune compared to the Abies and Picea species that dominate the premium market.
The species has been extensively used in dendrochronology. Many chronologies representing both subspecies exist. Studies have addressed climate reconstruction, archeological dating, ecological studies and stable isotope work supporting both climate and ecophysiological analysis.
Observations
Easily seen in the western USA and Canada. See subspecies menziesii and glauca for details.
Remarks
Douglas-fir was first described by Archibald Menzies (1754-1842), who discovered it on Vancouver Island during the 1790 voyage by Captain Vancouver to the northeast Pacific Ocean. The species' epithet honors Menzies. The name Douglas-fir commemorates David Douglas, who collected the species along the lower Columbia River in 1824; the seeds that he sent back to England were eagerly received and quickly established the tree as a garden favorite and an important agroforestry species.
"Douglas-fir ... has been a major component of the forests of western North America since the mid-Pleistocene (Hermann 1985). Although the fossil record indicates that the native range of Douglas-fir has never extended beyond western North America, the species has been successfully introduced in the last 100 years into many regions of the temperate forest zone (Hermann 1987)" (Lipscomb 1993).
Douglas-fir is the state tree of Oregon (Lipscomb 1993).
Both subspecies are host to the dwarf mistletoe Arceuthobium douglasii (Hawksworth and Wiens 1996).
This is one of the most-studied conifers in the world.
Citations
Adams, Robert P., J. Jesús Vargas-Hernández, M. Socorro González Elizondo, G. Hunter, T. A. Fairhall, David Thornburg, and Frank Callahan. 2012. Geographic variation in leaf essential oils of Douglas fir (Pseudotsuga menziesii). Phytologia 94(2).
Adams, Robert P., and M. Stoehr. 2013. Multivariate detection of hybridization using conifer terpenes II: Analyses of terpene inheritance patterns in Pseudotsuga menziesii F1 hybrids. Phytologia 95: 42–57.
Adams, R. P., J. Jesús Vargas-Hernández, M. Socorro González Elizondo, G. Hunter, T. A. Fairhall, David Thornburg, and Frank Callahan. 2013. Taxonomy of Douglas fir (Pseudotsuga menziesii) infraspecific taxa: vars. menziesii, glauca and oaxacana: nrDNA, cpDNA sequences and leaf essential oils. Phytologia 95(1).
Da Ronch, F., Caudullo, G., de Rigo, D., 2016. Pseudotsuga menziesii in Europe: distribution, habitat, usage and threats. In: San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A. (Eds.), European Atlas of Forest Tree Species. Publ. Off. EU, Luxembourg, pp. e01a4f5+
Davis, Owen K. 1999. Pollen Grain Morphology Larch. http://geo.arizona.edu/palynology/pid00013.html, accessed 2001.12.11, now defunct.
Flous, F. 1934a. Deux espèces nouvelles de Pseudotsuga Américains. Bull. Soc. Hist. Nat. Toulouse 66: 211-224.
Flous, F. 1934b. Diagnoses d'espèces et variétés nouvelles de Pseudotsuga Américains. Bull. Soc. Hist. Nat. Toulouse 66: 329-346.
Franco, J. do A. 1950. De coniferarum duarum nominibus. Boletin de la Sociedad Broteriana ser. 2 24:73-77.
Hermann, R. K. 1982. The genus Pseudotsuga: historical records and nomenclature. Oregon State University Forest Research Laboratory, Corvallis, OR.
Little, E.L., Jr. 1952. The genus Pseudotsuga (Douglas-fir) in North America. Leaflets Western Botany 6: 181-198.
Monumental Trees. 2006. Coast Douglas-fir in Goldkaul in Consdorf. https://www.monumentaltrees.com/en/lux/luxembourg/consdorf/5802_goldkaul/, accessed 2017.09.03.
Monumental Trees. 2013a. Coast Douglas-fir at Powerscourt House & Gardens. https://www.monumentaltrees.com/en/irl/leinster/wicklow/6729_powerscourthousegardens/15733/, accessed 2017.09.03.
Monumental Trees. 2013b. Coast Douglas-fir in the arboretum Saint-Michel. https://www.monumentaltrees.com/en/bel/luxembourg/sainthubert/492_arboretumsaintmichel/15992/, accessed 2017.09.03.
Christiansen, Per. 2018.05.27. Ny rekord: Nordens højeste træ står i Østjylland. TV2 Østjylland. https://www.tv2ostjylland.dk/artikel/partner/tv2dk/ny-rekord-nordens-hoejeste-trae-staar-i-oestjylland, accessed 2019.07.22 (includes video of the climbers making the measurement).
Monumental Trees. 2017a. Coast Douglas-fir 'Highest tree in Czech republic' near the Vlastibor in Zelezný Brod. https://www.monumentaltrees.com/en/cze/liberecky/jablonecnadnisou/17056_vlastibor/, accessed 2017.09.03.
Monumental Trees. 2017b. Coast Douglas-fir at the royal estate Het Loo. https://www.monumentaltrees.com/en/nld/gelderland/apeldoorn/209_royalestatehetloo/3028/, accessed 2017.09.03.
Oswalt, S. N., W. B. Smith, P. D. Miles, and S. A. Pugh. 2019. Forest resources of the United States, 2017. Gen. Tech. Rep. WO-97. Washington, DC: U.S. Department of Agriculture, Forest Service, Washington
Office. 223 p. https://doi.org/10.2737/WO-GTR-97.
Peng Li and W.T. Adams. 1989. Rangewide patterns of allozyme variation in Douglas-fir. Canadian Journal of Forest Research 19: 149-161.
Pulkrab, K., M. Sloup, and M. Zeman. 2014. Economic Impact of Douglas-fir (Pseudotsuga menziesii
[Mirb.] Franco) production in the Czech Republic. Journal of Forest Science 60(7):297-306.
Reveal, James R. [no date]. A Nomenclatural Morass. http://www.plantsystematics.org/reveal/pbio/LnC/dougfir.html, accessed 2011.05.20.
Schmid, M., M. Pautasso, and O. Holdenrieder. 2014. Ecological consequences of Douglas fir (Pseudotsuga menziesii) cultivation in Europe. European Journal of Forest Research, 133: 13–29.
Snajberk, K. and E. Zavarin. 1976. Mono- and sesqui-terpenoid differentiation of Pseudotsuga of the United States and Canada. Biochem. Syst. Ecol. 4:159-163.
Van Pelt, R., and Sillett, S. C. 2008. Crown development of coastal Pseudotsuga menziesii, including a conceptual model for tall conifers. Ecological Monographs 78(2):283–311.
Venturini, Glenda. 2016.12.28. L'albero più alto d'Italia? È a Vallombrosa: scalato e misurato da un team di esperti. http://valdarnopost.it/news/l-albero-piu-alto-d-italia-e-a-vallombrosa-scalato-e-misurato-da-un-team-di-esperti, accessed 2017.09.03.
See also
Arno, Stephen F. and Jane Gyer. 1973. Discovering Sierra trees. Yosemite Natural History Association. 89pp.
Arno, Stephen F. and Carl E. Fiedler. 2020. Douglas fir the story of the West's most remarkable tree. Seattle: Mountaineers Books. 192 pp.
Elwes and Henry 1906-1913 at the Biodiversity Heritage Library (Photos). This series of volumes, privately printed, provides some of the most engaging descriptions of conifers ever published. Although they only treat species cultivated in the U.K. and Ireland, and the taxonomy is a bit dated, still these accounts are thorough, treating such topics as species description, range, varieties, exceptionally old or tall specimens, remarkable trees, and cultivation. Despite being over a century old, they are generally accurate, and are illustrated with some remarkable photographs and lithographs.
FEIS database.
Lanner (1983).
Sargent (1898) provides an exceptionally detailed description of this species (using the name Pseudotsuga mucronata), with an excellent illustration.